致病性τ减少核紧张局势培养的神经元
 Claira孙
Claira孙 嘉成马
嘉成马 威廉·j·雷
威廉·j·雷 贝丝霜
贝丝霜- 1电池系统和解剖学、Barshop长寿和衰老研究所格伦·比格斯阿尔茨海默氏症和神经退行性疾病研究所,德克萨斯大学健康圣安东尼奥马刺,TX,美国
- 2神经退化财团,治疗发现,德克萨斯大学MD安德森癌症中心,休斯顿,美国TX
神经退行性tauopathies,包括阿尔茨海默病病理上定义的聚合形式的τ蛋白影响人的大脑。先前的研究报告,致病性τ的负面影响和肌动蛋白细胞骨架微管导致有毒的不稳定的核纤层蛋白nucleoskeleton和核内陷和气泡的形成。基于核的已知函数mechanosensor,以及核多形性的高发病率在人类阿尔茨海默病及相关tauopathies,我们研究了致病性τ核紧张局势的影响。我们第一次发现tau-dependent核膜内陷和relocalization Nucleoskeleton链接器和细胞骨架(林肯)复杂组件保存在一个新近发展起来的神经母细胞瘤细胞系,doxycycline-inducible表达式τ突变与常染色体显性遗传的额颞叶痴呆。我们下决定,福斯特共振能量转移(FRET)的核张力传感器响应细胞骨架稳定和不稳定表达在神经母细胞瘤细胞。使用这个核张力传感器,我们发现致病的诱导表达τ足以减少核紧张。这项工作提供了最初的概念验证的证据,致病形式的τ改变核紧张,为未来铺平道路的上下文中研究改变核mechanosensing tau-mediated神经退行性疾病。
1介绍
人类的身体经历了持续的机械应力。心脏的每个脚步或击败,细胞拉伸和压扁保持器官的完整性和功能。虽然是一个不动的器官保护的头骨,大脑也受到机械应力。一个广泛的大脑血管促进不断的血液在大脑和维持血脑屏障。大脑血管直接与细胞的相互作用,使每个细胞经历了不同水平的机械应力。例如,在老鼠、躯体感觉刺激单一软膜的小动脉的直径增加30% (Ngai,韦恩,1996)。物离子通道的存在如TRAAK (Maingret et al ., 1999)和TREK-1 (Hervieu et al ., 2001)在神经元进一步表明,这些细胞的质膜正准备应对机械信号的变化而导致的血管的直径的差异导致血液流动。事实上,研究表明,机械刺激诱导培养的神经元神经钙响应(Gaub et al ., 2020)。
而很明显,细胞经历机械应力和机械应力可以改变在生理和病理条件下,相对较少的已知关于的mechanosensing能力在大脑细胞(Kirby Lammerding, 2018)。在边缘,细胞力学生物学的一个重要的作品已经确定细胞细胞核作为关键的探测器和应答器机械应力(Kirby Lammerding, 2018;毛雷尔Lammerding, 2019;Hamouda et al ., 2020;Janota et al ., 2020)。最近的研究在人类上皮细胞祖细胞表明原子核发生机械软化,防止机械应激损伤(纳瓦et al ., 2020)。机械信号传达从胞质细胞核(反之亦然)通过核envelope-spanning林肯复杂(图1)。在林肯复杂,lamin-bound太阳蛋白质形成三聚物从核纤层蛋白中间丝在细胞核进入细胞核周围的空间直接绑定到nesprin蛋白质。Nesprins从细胞核周围的空间延伸到细胞质中,在那里他们可以直接绑定到肌动蛋白丝,与微管马达,并将间接绑定到中间丝(Janota et al ., 2020)。虽然mechanosensing林肯复杂的属性尚未研究神经元,研究在人类间充质干细胞主要报告说,林肯复杂紧张直接影响细胞的形状,染色质压实、转录后修饰、蛋白质组(吉尔伯特et al ., 2019)。

图1。林肯复杂,其表示细胞和nucleoskeleton直接交互。
一个积累的作品表明,核纤层蛋白nucleoskeleton经历扰动和凹入/出泡细胞窝藏致病形式的τ(弗罗斯特et al ., 2016;Montalbano et al ., 2019;Paonessa et al ., 2019;江和Wolozin, 2021)。野生型和突变体形式的τ蛋白可以组装成可溶性有毒multimeric物种,大型神经原纤维缠结和夹杂物,可以驱动神经退化果蝇接受疾病有关的转译后的修改和构象的变化没有探测到multimerization (维特曼et al ., 2001;Steinhilb et al ., 2007),我们定义“致病”形式的τ与携带phosphoepitopes与人类疾病相关的当前研究的目的。与整体减少大脑的核纤层蛋白的蛋白质果蝇tauopathy模型和人类大脑受到阿尔茨海默氏症、神经核特性普遍decondensation异染色质和广泛的核内陷和起泡。在果蝇,基因操作的肌动蛋白聚合和林肯复杂的组件协调tau-induced神经退化和核架构的变化,表明之间的因果联系τ的肌动蛋白细胞骨架的影响,林肯复杂、核纤层蛋白nucleoskeleton和神经退行性变的(弗罗斯特et al ., 2016)。最近的研究在培养细胞和初级老鼠组织表明,核被膜完整性的破坏可能是启动事件tauopathies (Prissette et al ., 2022)。虽然有这样一个清晰的分解nucleoskeleton和形态变化的神经核的上下文中tauopathy,迄今为止没有一项研究调查了细胞核在细胞的力学性能窝藏致病形式的τ。
力学生物学研究涉及发展的一个基本的进步FRET-based张力传感器模块“TSmod”(Grashoff et al ., 2010)。TSmod特性一个弹性肽在mTFP1和金星荧光蛋白。力投入TSmod引起的弹性伸长肽,从而降低整个担心信号通过减少能源从mTFP1捐赠者转移到金星受体。TSmod生物传感器已经插入各种蛋白质,驻留在细胞间和cell-matrix粘附网站为了全面量化细胞张力(Grashoff et al ., 2010;Borghi et al ., 2012;康威et al ., 2013;Cai et al ., 2014;栗山et al ., 2014;奥斯丁et al ., 2015)。在当前的研究中,我们利用一个版本的TSmod插入功能Nesprin-2G构造之间的机械部队,允许量化肌动蛋白细胞骨架和核被膜(Arsenovic康威,2018)。近年来,这个nesprin张力传感器(nesprin-TS)已经被用来测量机械力在林肯复杂的成纤维细胞,胚胎干细胞,myofibroblasts和上皮细胞(Arsenovic康威,2018;胡锦涛等人。,2019年;沃克et al ., 2021;Pothapragada et al ., 2022)。
我们结合一个诱导模型的tauopathy nesprin张力传感器发现致病τ的诱导表达导致培养全面减少核紧张神经母细胞瘤细胞。这些结果强调了mechanosensing属性核的神经元和为未来奠定了基础调查致病性的影响形式的核转导τ。
2材料和方法
2.1互补脱氧核糖核酸结构和病毒
MAPT互补脱氧核糖核酸(NM_005910.6)单一突变(1216 c > T)和绿色荧光蛋白互补脱氧核糖核酸合成,插入到doxycycline-inducible慢病毒载体PLIX_403 (Addgene # 41395)。慢病毒粒子生成的MD安德森功能基因组学的核心设备。
2.2细胞培养和iTau建立稳定的细胞系
人类神经母细胞瘤细胞((2)- c;写明ATCC # crl - 2268)在1:1的混合培养鹰的最低基本培养基(EMEM)和F12培养基补充penicillin-streptomycin tetracycline-free的边后卫为10%和1%。稳定细胞池是(2)-C_MAPT-R406W和(2)-C_GFP转换生成的是(2)- c细胞PLIX_403_MAPT-R406W或PLIX_403_GFP慢病毒粒子,其次是选择2μg /毫升嘌呤霉素。MAPT-R406W或GFP的表达与1μg /毫升强力霉素诱导hyclate溶解在DMSO溶液。
细胞被镀在不需抗生素OptiMEM(低血清媒体)(ThermoFisher # 31985062)转染前24小时密度允许80%∼-85% confluency第二天。细胞转染nesprin-TS或nesprin-HL(分别Addgene # 68127和# 68128)使用Lipofectamine 2000 (ThermoFisher, # 11668030)。1µg DNA和2.5µL Lipofectamine稀释96.5µL OptiMEM每个12-well板。鸡尾酒是在室温下孵化25分钟前增加了细胞。细胞被孵化与DNA / Lipofectamine OptiMEM 6小时前媒体替换。收集细胞转染48 h后各自的实验。
对强力霉素诱导转基因表达,细胞被镀在媒体不低血清(2.5% tetracycline-free的边后卫EMEM补充)24小时前密度允许85%∼-90% confluency第二天。细胞孵育1μg /毫升的强力霉素或车辆(DMSO)大约24小时前集合。
2.3免疫印迹
细胞收获∼90% confluency用里帕1 x蛋白酶抑制剂(停止蛋白酶,ThermoFisher),然后在4°C的环境与温柔摇30分钟。细胞溶解产物离心20分钟在4°C 12000 rpm。一个布拉德福德蛋白质含量测定细胞上清液进行量化(BCA蛋白质化验设备,皮尔斯)之前西方墨点法。蛋白溶解产物中煮2 x Laemmli缓冲5分钟,在12000转离心1分钟,然后加载到4% - -20% sds - page凝胶。20μg蛋白质是加载/。等于负载评估了朱红色年代染色后硝化纤维膜的转移。膜被封锁在PBS包含0.05%的渐变和2%的牛奶,然后孵化主要抗体在一夜之间在4°C。洗后,膜被孵化与HRP-conjugated二级抗体在室温下2 h。墨迹是增强化学发光底物。使用ImageJ微了。
2.4免疫荧光
细胞被镀在20毫米12-well盘子盖玻片前染色。细胞被固定在冰冷的分子级100%甲醇室温5分钟,然后洗了三次有0.01%在PBS Tween-20每洗5分钟。洗后,细胞permeabilized 0.01% PBS Triton x - 100年有1%的牛血清白蛋白(BSA)在室温下15分钟,用PBS Tween-20 0.01%洗了三次,然后封锁了30分钟的PBS Tween-20 BSA为1%和0.01%。主要抗体在一夜之间被稀释1% BSA和孵化细胞在4°C。第二天,细胞用PBS Tween-20 0.01%,然后用二次孵化抗体室温1小时。洗后,细胞染色2分钟1 x DAPI染色细胞核,然后安装到玻璃幻灯片Vectashield (# h - 1000 - 10, Vectorlabs)。Latrunculin经处理的细胞被孵化Acti-Stain 555 Phalloidin (# PHDH1-A,细胞骨架)之前DAPI染色。用共焦显微镜观察细胞(710年蔡司LSM NLO考官或蔡司LSM 880)。ImageJ用于分析。
计算比例的细胞核含有核内陷,100细胞/复制得分内陷或气泡的存在。使用ImageJ核Nesprin-1荧光进行量化。核面具创建基于DAPI染色,平均强度Nesprin-1面具内计算为每个复制。
2.5烦恼成像和分析
Pre-bleach和post-bleach图像获得使用蔡司LSM 880共焦显微镜。收购pre-bleach图像后,受体(金星)photobleached波长515纳米的激光线的100%力量200次迭代。第二张照片是光漂白后获得的。荧光强度是通过选择一个感兴趣的区域在核膜。担心效率计算,减去捐赠(mTFP1)强度从捐献者光漂白后强度光褪色之前,然后除以光漂白后的捐赠。每个数据点代表烦恼分析在一个核。三十核分析了在三个不同的井(10核/)。所有烦恼分析单个实验进行当天在会议一个显微镜。
3的结果
3.1τ的诱导表达R406W导致核内陷和再分配的林肯在培养的神经母细胞瘤细胞复杂的蛋白质
神经核窝藏致病形式的τ的发生率显著增加起泡和内陷(弗罗斯特et al ., 2016;Montalbano et al ., 2019;Paonessa et al ., 2019;江和Wolozin, 2021)。研究果蝇tauopathy模型表明,核膜内陷和气泡的负面影响的结果致病形式的τ的肌动蛋白细胞骨架和林肯复杂。除了核多形性人类阿尔茨海默氏症的大脑和τ转基因果蝇,神经细胞核受到τ有b型核纤层蛋白的整体水平降低(弗罗斯特et al ., 2016),蛋白质形成中间丝,行神经核的内部表面建立核结构和强度(至关重要Worman 2012)。
作为第一步确定致病性的影响τ核结构改变核紧张,我们开发了一个(2)- c神经母细胞瘤细胞模型特性doxycycline-inducible表达人类τ携带额颞叶dementia-associated突变R406W(赫顿et al ., 1998)(τR406W)。我们这个模型”一词iTau。“我们发现健壮的表达人类疾病相关τphosphoepitopes iTau细胞24小时后与强力霉素诱导车辆相比治疗控制(图2一个;补充图S1A)。作为第二控制,我们证实细胞doxycycline-induced GFP的表达,“iGFP”,不累积变异phosphotau (补充图印地)。报道在人类阿尔茨海默病,τ转基因果蝇(弗罗斯特et al ., 2016;Cornelison et al ., 2019),诱导多能干细胞(iPSC)中神经元窝藏变异τ(MAPT)突变(Paonessa et al ., 2019),主要与诱导神经元τ聚合(江和Wolozin, 2021)我们发现致病的诱导表达τ足以形成核膜内陷和气泡iTau细胞(图2 b)。我们不能独自观察强力霉素治疗的作用是(2)- c核形态(补充图就是S1C)。

图2。诱导致病形式的τiTau细胞导致核膜内陷和再分配林肯复杂的组件。(一)检测抵抗τphosphoepitopes iTau细胞在强力霉素治疗基于西方墨点法。(B)24小时doxycycline-mediated感应τ表达会导致显著增加核膜内陷车辆治疗控制相比,基于核纤层蛋白B1免疫荧光。(C)SUN1关系(C)和SUN2(D)与核内陷和气泡在doxycycline-mediated感应τiTau细胞中表达。空的箭头(D)表明存在SUN2核气泡核纤层蛋白缺乏。(E)Doxycyline-mediated诱导τiTau细胞表达导致显著增加核Nesprin-1基于免疫荧光。n= 3复制/组,t以及,*p< 0.05,* *p< 0.01。误差线表明SEM。
以前的工作报告,林肯复杂有原因地调和τ核架构的影响,和太阳本地化核膜在τ转基因改变果蝇(弗罗斯特et al ., 2016),我们接下来分析了林肯的定位复杂组件SUN1 SUN2, Nesprin-1 doxycycline-inducedτ后iTau细胞表达。我们观察到SUN1 tau-induced核膜内陷和气泡iTau细胞(图2 c)。SUN2遵循类似的模式,与罕见的实例中气泡对SUN2是有利的,但缺乏核纤层蛋白(图2 d)。我们检测到显著增加核本地化的Nesprin-1诱导τ表达式(图2 e)。我们下一个决心如果诱导τ表达式改变整体水平的林肯复杂和核纤层蛋白B1蛋白质含量。基于免疫印迹,我们没有发现显著差异在SUN1 SUN2, Nesprin-1,或核纤层蛋白B1蛋白质水平doxycycline-mediated诱导τ表达式(补充数据S2A-C)。这些初步分析支持以前的工作报告,致病形式的τ改变林肯复杂的定位和驱动核内陷,形成气泡在其他模型系统和人类tauopathy (弗罗斯特et al ., 2016)。
3.2开发核张力传感器在神经母细胞瘤细胞
林肯复杂组件Nesprin-2巨头(Nesprin-2G)是一个大型800 kD蛋白港口两个actin-binding域N末端,紧随其后的是一长段spectrin-like重复,一个跨膜域和制造商域与太阳相互作用蛋白质在核膜的双分子层(Padmakumar et al ., 2004)。nesprin-TS构造由TSmod插入Nesprin-2G的微型版本,“mini-nesprin-2G”,缺乏大部分的血影蛋白重复(Arsenovic康威,2018)。担心捐赠(mTFP1)和烦恼受体(金星)由一个HP35弹性肽(奥斯丁et al ., 2015)。在这个系统中,FRET-based nesprin-TS信号是成反比的肌动蛋白细胞骨架之间的紧张关系和核纤层蛋白nucleoskeleton。控制,我们利用nesprin无头(nesprin-HL)的一个版本nesprin-TS构造缺乏肌动蛋白结合域,因此对部队(Arsenovic康威,2018)(图3一)。转染后48 h的(2)- c细胞nesprin-TS生物传感器和nesprin-HL控制,我们进行受体photobleach-based烦恼分析。我们发现显著降低焦虑水平相比nesprin-TS nesprin-HL (图3 b),确认nesprin-TS生物传感器功能设计在这个细胞模型。
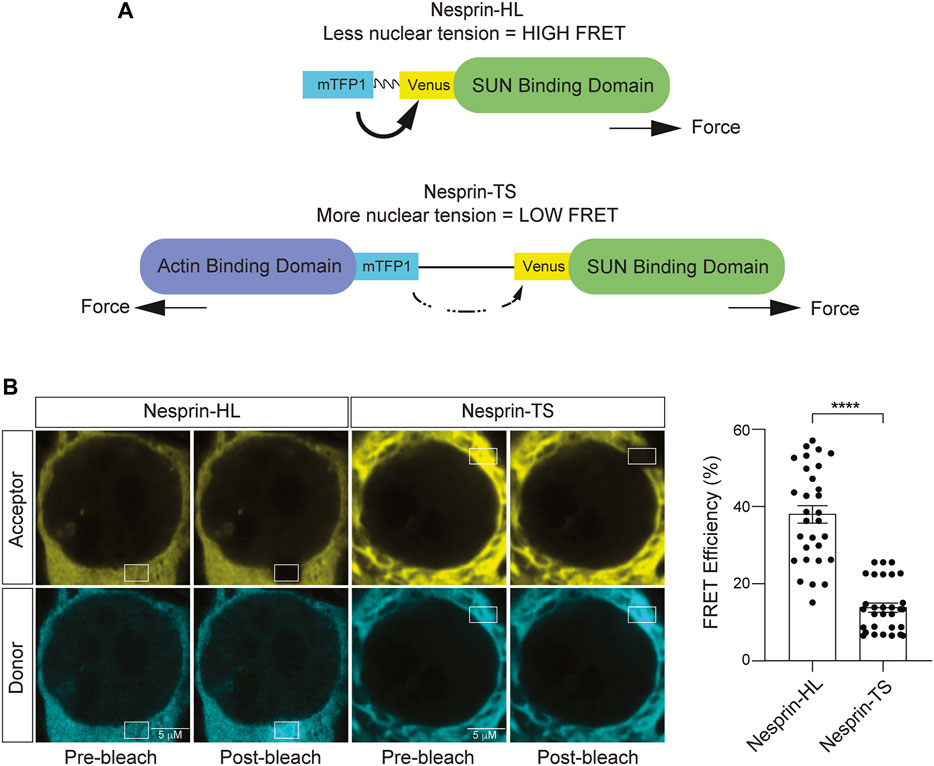
图3。神经母细胞瘤细胞中核张力传感器的发展。(一)示意图nesprin-HL nesprin-TS和逆烦恼和核之间的关系紧张。(B)受体photobleach-based量化的烦恼nesprin-HL和nesprin-TS表达(2)- c细胞。每个数据点代表烦恼分析在一个核。三十核分析了在三个不同的井(10核/),t以及,* *p< 0.01。误差线表明SEM。
3.3 Latrunculin A-induced损耗的丝状肌动蛋白减少核紧张神经母细胞瘤细胞
在成功表示nesprin-TS是(2)- c细胞和量化核紧张通过烦恼,我们接下来问nesprin-TS回应核紧张由肌动蛋白细胞骨架的变化。以前的研究报告说,latrunculin抑制丝状肌动蛋白的形成(f -肌动蛋白)(斯佩克特et al ., 1983)。我们发现下半场治疗神经母细胞瘤细胞的10µM latrunculin足以有效地消耗总体水平的f -肌动蛋白基于phalloidin染色(图4一),latrunculin一个调解的f -肌动蛋白不稳定引起浅凹入的核纤层蛋白nucleoskeleton (图4 b)。Latrunculin治疗增加烦恼的效率(2)- c细胞窝藏nesprin-TS (图4 c),这表明f -肌动蛋白不稳定足以减少核紧张局势培养神经母细胞瘤细胞。

图4。Latrunculin A-induced损耗f -肌动蛋白减少核紧张局势神经母细胞瘤细胞。(一)Phalloidin-based可视化的f -肌动蛋白(2)- c细胞治疗10µM latrunculin或车辆(DMSO) 15分钟。(B)Latrunculin治疗驱动器增加核膜内陷检测到核纤层蛋白B1免疫荧光。n= 3复制/组,t以及,*p< 0.05。(C)受体photobleach-based量化的烦恼nesprin-TS表达(2)- c细胞在治疗10µM latrunculin汽车相比。每个数据点代表烦恼分析在一个核。三十核分析了在三个不同的井(10核/),t以及,* * * *p< 0.0001。误差线表明SEM。
3.4 Jasplakinolide-induced overstabilization丝状肌动蛋白增加核紧张局势神经母细胞瘤细胞
我们接下来问增加细胞骨架刚度影响核紧张局势(2)- c细胞。over-stabilize肌动蛋白细胞骨架,我们利用jasplakinolide cyclo-depsipeptide绑定和稳定肌动蛋白二聚体,从而提高成核的f -肌动蛋白(Bubb et al ., 1994)。作为jasplakinolide竞争性抑制绑定phalloidin f -肌动蛋白,我们可视化jasplakinolide-treated细胞的肌动蛋白细胞骨架通过染色的细胞检测的抗体β肌动蛋白。我们观察到明显的海拔β1小时后肌动蛋白治疗与0.5 (2)- c细胞µM jasplakinolide (图5一个)。后jasplakinolide-mediated overstabilization肌动蛋白细胞骨架,我们发现核膜内陷的增加(图5 b)。使用nesprin-TS生物传感器,我们发现f -肌动蛋白overstabilization显著减少烦恼的细胞是(2)- c (图5 c),这表明过于刚性骨架增加核紧张局势这个模型。在某些情况下,核紧张的程度是如此之高,以至于担心效率有效零,表明荧光团作为一对nesprin-TS不再工作。

图5。Jasplakinolide-induced海拔f -肌动蛋白增加核紧张神经母细胞瘤细胞。(一)Immunofluorescence-based 1小时治疗后肌动蛋白细胞骨架的检测与0.5 (2)- c细胞µM jasplakinolide或车辆(DMSO)。(B)Jasplakinolide治疗诱发显著增加核膜内陷基于核纤层蛋白B1免疫荧光。n= 3复制/组,t以及,*p< 0.05。(C)受体photobleach-based量化的烦恼nesprin-TS表达与0.5 (2)- c细胞在治疗µM jasplakinolide相比。每个数据点代表烦恼分析在一个核。三十核分析了在三个不同的井(10核/),t以及,*p< 0.05,* * * *p< 0.0001。误差线表明SEM。
3.5表达式的致病性τ导致核减少紧张
建立核内陷和起泡,iTau细胞特性,随着relocalization林肯复杂的组件,这nesprin-TS忠实地检测诱导核紧张局势的变化,我们接下来问核张力改变的环境中致病τ。作为强力霉素的潜在影响的控制核紧张,我们首先确定如果强力霉素治疗影响核紧张局势没有诱导τ表达式。我们发现24 h(强力霉素治疗显著减少烦恼,表明增加核紧张神经母细胞瘤细胞(图6)。这个警告,我们接下来介绍nesprin-TS iTau细胞和量化核紧张通过烦恼。尽管强力霉素的能力降低整体烦恼没有τ,我们发现doxycycline-mediated诱导致病τ表达显著增加烦恼,揭示一个健壮的τ的能力来有效地减少核紧张(图6 b)。

图6。诱导τiTau细胞表达导致减少核紧张。(一)受体photobleach-based量化烦恼的是(2)- c细胞强力霉素治疗24小时后显示doxycycline-induced增加核紧张相比,车辆(DMSO)细胞治疗。(B)受体photobleach-based量化iTau细胞24 h后的烦恼doxycycline-mediated感应τ表达相比,车辆显示tau-induced降低核紧张。每个数据点代表烦恼分析在一个核。三十核分析了在三个不同的井(10核/),t以及,* *p< 0.01。误差线表明SEM。
4讨论
致病形式的τ与核多型现象在各种实验室模型和人类疾病(弗罗斯特et al ., 2016;Montalbano et al ., 2019;Paonessa et al ., 2019;江和Wolozin, 2021)。提出了几种non-mutually排斥机制来解释τ对原子核的三维体系结构的影响。研究果蝇表明tau-induced肌动蛋白overstabilization驱动器relocalization林肯复杂的蛋白质,调解一个不稳定的核纤层蛋白nucleoskeleton, decondensation结构异染色质,顺向神经退化(弗罗斯特et al ., 2016)。符合这些发现,核内陷神经元从人类阿尔茨海默病大脑组织被发现港口核心的丝状肌动蛋白的蛋白质。iPSC-derived神经元的研究报告,τ的负面影响在微管动力学驱动器核膜内陷,,核膜内陷包含聚合的微管蛋白(Paonessa et al ., 2019)。这两个研究发现致病形式的τ内存在核膜内陷。最近的一项研究在初级神经元报道optogenetic诱导τ寡聚化变形核膜在几分钟内,导致的直接交互寡聚τ物种与核纤层蛋白和核孔蛋白(江和Wolozin, 2021)。基于这些研究,以及已知的肌动蛋白参与,林肯复杂,核纤层蛋白,和异染色质的核监管机构刚度,和2)传播和响应的机械信号和核(Janota et al ., 2020;纳瓦et al ., 2020;沃克et al ., 2021),我们被问及细胞核的细胞受到致病性τ接受核紧张局势的变化。
与以前的工作相一致果蝇,从病人窝藏iPSC-derived神经元MAPT与诱导突变,主要培养皮层神经元τ齐聚反应,诱导τ在HEK293细胞中表达,和后期人类阿尔茨海默病大脑组织(弗罗斯特et al ., 2016;Montalbano et al ., 2019;Paonessa et al ., 2019;江和Wolozin, 2021),我们观察到显著增加核内陷和起泡的培养和诱导神经母细胞瘤细胞τ表达相比,控制。此外,我们发现太阳蛋白质tau-induced核内陷和气泡,均有密切的联系,整体水平的核Nesprin-1升高tau-expressing神经母细胞瘤细胞。符合我们发现原子核受到致病性τ功能核内陷和起泡的发病率高与relocalization林肯复杂的组件(弗罗斯特et al ., 2016),我们发现病理τ的诱导表达减少核紧张。
虽然上面引述的研究清楚地表明,细胞因子调节核紧张tauopathy中特异表达,我们没有集中在一个特定的候选人机制推动tau-induced减少核紧张在当前的研究中。致病性τ是报道引起肌动蛋白细胞骨架的overstabilization (Fulga et al ., 2007;他et al ., 2009年;卡布拉尔Fontela et al ., 2017),我们认为也许这致病性τ增加刚度的核基于我们发现jasplakinolide-induced f -肌动蛋白浓缩足以增加核紧张局势。另一方面,减少核纤层蛋白B1和核软化肺癌和乳腺癌的发生在细胞培养模型(贾et al ., 2019;费舍尔et al ., 2020),和淘汰赛核纤层蛋白A / C或诱导异染色质decondensation都足以减少核刚度(Lammerding et al ., 2004;Lammerding et al ., 2006;纳瓦et al ., 2020)。据报道,而核纤层蛋白B1缺乏推动核起泡没有改变核刚度在小鼠胚胎成纤维细胞(Lammerding et al ., 2006),超表达的核纤层蛋白B1 HEK293核刚度和N2a细胞增加,以及患者的细胞基因复制的基因编码核纤层蛋白B1 (费雷拉et al ., 2014)。基于这些研究,我们推测,减少核紧张我们观察神经母细胞瘤细胞诱导τ表达式是nucleoskeleton政局不稳的结果(弗罗斯特et al ., 2016;Montalbano et al ., 2019)和结构异染色质decondensation (弗罗斯特et al ., 2014)。
由于其直接互动与肌动蛋白细胞骨架和核纤层蛋白nucleoskeleton,林肯复杂的检测和传送机械信息与原子核(Janota et al ., 2020)。据报道,增加核紧张诱发核纤层蛋白A / C的招聘到林肯复杂(Guilluy et al ., 2014),这表明机械拉动nesprins可能触发的强化核纤层蛋白nucleoskeleton。研究皮肤表皮祖细胞提供洞察生物的好处核张力变化,由于外力(纳瓦et al ., 2020)。在这些研究中,发现核软化响应机械力来保护细胞对DNA损伤的积累产生的反应细胞伸展。目前我们不知道tau-induced核软化有原因地介导神经元死亡,有可能减少核紧张可以作为补偿响应tauopathy保护DNA免受机械及外力损伤。虽然我们的数据表明,变异的诱导表达MAPT突变τ核紧张,造成损失的,重要的是要注意,目前我们不知道生理形式的τ也在十几调节细胞核的机械性能设置。
除了作为mechanosensor (Kirby Lammerding, 2018),细胞核作为mechanoresponsive细胞器,促进细胞反应的改变机械环境。核适应机械应力包括一系列生物过程,包括微分表达式“mechanosensitive基因,蛋白质转译后的修改和本地化,核孔隙渗透率和mechanosensitive离子通道的变化,和细胞谱系的决心(毛雷尔Lammerding, 2019)。增加核孔的“泄漏”(Eftekharzadeh et al ., 2018)和海拔的RNA出口tauopathy模型(Cornelison et al ., 2019)符合改变核转导核孔的影响。同样的,我们曾报道,iPSC-derived神经元的神经细胞核从人类以及τ转基因阿尔茨海默氏症病例果蝇功能衰竭的核钙(Mahoney et al ., 2020)。虽然这些细胞表型与致病的潜在影响是一致的形式的核转导τ,仍有待确定tau-induced核刚度变化的直接驱动tauopathy核钙变化和核孔隙渗透率。
虽然我们的研究是第一个研究神经元核紧张局势并提供第一个洞察致病性τ核力学的影响,我们注意到一些我们的方法的局限性。人类的大脑是一个,而软组织与杨氏模量从1.389∼-1.895 kPa (Budday et al ., 2015),神经元生长在玻璃盖玻片体验一种明显不同的环境比人类大脑中的神经元。此外,大脑血管常数所产生的机械应力并不存在于我们的系统,不同的细胞类型,也构成了生活的大脑。随着各种癌症与核紧张局势的变化,我们使用一个永生化细胞系可能导致τ核紧张局势的影响。最后,虽然我们推测,我们的发现并不局限于MAPT R406W突变,零星的人类tauopathy和各种型号的tauopathy野生型形式的τ也表现出核多形性,未来的研究需要确定毒性形式的野生型τ诱导核软化。
当前的理解核转导几乎完全基于non-neuronal细胞类型。我们的研究结果提供理论水平的数据,为未来研究提供理论基础核转导τ的影响和发展的系统来更好地学习核神经元力学如iPSC-derived脑瀑样或更多的生理环境在活的有机体内模型。与知识的大脑细胞的确受到机械应力,我们的分析提供了第一个见解的反应神经核外力的肌动蛋白细胞骨架,以及核紧张局势发生变化由于致病性τ。
数据可用性声明
最初的贡献提出了研究中都包含在这篇文章/补充材料,进一步的调查可以针对相应的作者。
作者的贡献
实验设计、写作、和图是由CS和男朋友做准备。通过计算机科学实验和数据分析。iTau / iGFP细胞系是由JM WJR。
资金
这项研究得到了国家老龄研究所[R01 AG057896 (BF) T32 AG021890 (CS)]和国家综合医学科学研究所R25 GM095480 (CS)。WR和JM神经退化财团支持的贝尔弗家族基金会和奥斯卡·费舍尔项目。
确认
我们感谢丹尼尔·康威提供洞察力和建议使用nesprin-TS和nesprin-HL传感器,和吉姆Lechleiter和前男友王建议受体photobleaching-based量化的烦恼。显微镜的一部分进行了圣安东尼奥的UT健康核心光学成像设备,这是由NIH-NCI P30 CA54174。图1与BioRender创建。
的利益冲突
作者声明,这项研究是在没有进行任何商业或财务关系可能被视为一个潜在的利益冲突。
出版商的注意
本文表达的所有索赔仅代表作者,不一定代表的附属组织,或出版商、编辑和审稿人。任何产品,可以评估在这篇文章中,或声称,可能是由其制造商,不保证或认可的出版商。
补充材料
本文的补充材料在网上可以找到:https://www.雷竞技rebatfrontiersin.org/articles/10.3389/fragi.2023.1058968/full补充材料
引用
Arsenovic, p . T。,和康威,d E。(2018). Using nesprin tension sensors to measure force on the LINC complex.摩尔。生物方法。1840年,59 - 71。doi: 10.1007 / 978 - 1 - 4939 - 8691 - 0 - _6
奥斯丁,K。,Ringer, P., Mehlich, A., Chrostek-Grashoff, A., Kluger, C., Klingner, C., et al. (2015). Extracellular rigidity sensing by talin isoform-specific mechanical linkages.Nat,细胞生物。17日,1597 - 1606。doi: 10.1038 / ncb3268
Borghi, N。,Sorokina, M., Shcherbakova, O. G., Weis, W. I., Pruitt, B. L., Nelson, W. J., et al. (2012). E-cadherin is under constitutive actomyosin-generated tension that is increased at cell–cell contacts upon externally applied stretch.Proc。国家的。学会科学。109年,12568 - 12573。doi: 10.1073 / pnas.1204390109
Bubb, m R。,Senderowicz, A. M., Sausville, E. A., Duncan, K. L., and Korn, E. D. (1994). Jasplakinolide, a cytotoxic natural product, induces actin polymerization and competitively inhibits the binding of phalloidin to F-actin.生物。化学。269年,14869 - 14871。doi: 10.1016 / s0021 - 9258 (17) 36545 - 6
Budday, S。,不,R。,de Rooij, R., Steinmann, P., Wyrobek, T., Ovaert, T. C., et al. (2015). Mechanical properties of gray and white matter brain tissue by indentation.j .机械工程。Behav。生物医学。母亲46岁,318 - 330。doi: 10.1016 / j.jmbbm.2015.02.024
卡布拉尔Fontela Y。Kadavath, H。Biernat, J。里德尔,D。,Mandelkow, E., and Zweckstetter, M. (2017). Multivalent cross-linking of actin filaments and microtubules through the microtubule-associated protein Tau.Commun Nat。8,1981。doi: 10.1038 / s41467 - 017 - 02230 - 8
Cai, D。,Chen, S.-C., Prasad, M., He, L., Wang, X., Choesmel-Cadamuro, V., et al. (2014). Mechanical feedback through E-cadherin promotes direction sensing during collective cell migration.细胞157年,1146 - 1159。doi: 10.1016 / j.cell.2014.03.045
康威,d E。,Breckenridge, M. T., Hinde, E., Gratton, E., Chen, C. S., and Schwartz, M. A. (2013). Fluid shear stress on endothelial cells modulates mechanical tension across VE-cadherin and PECAM-1.咕咕叫。医学杂志。23日,1024 - 1030。doi: 10.1016 / j.cub.2013.04.049
Cornelison, g . L。,征税,美国。简森,T。,和霜,B。(2019). Tau-induced nuclear envelope invagination causes a toxic accumulation of mRNA in果蝇。衰老细胞18日,e12847。doi: 10.1111 / acel.12847
Eftekharzadeh B。Daigle, j·G。,Kapinos, L. E., Coyne, A., Schiantarelli, J., Carlomagno, Y., et al. (2018). Tau protein disrupts nucleocytoplasmic transport in Alzheimer’s disease.神经元99年,925 - 940。e7。doi: 10.1016 / j.neuron.2018.07.039
费雷拉,D。,Canale, C., Marotta, R., Mazzaro, N., Gritti, M., Mazzanti, M., et al. (2014). Lamin B1 overexpression increases nuclear rigidity in autosomal dominant leukodystrophy fibroblasts.美国实验生物学学会联合会J。28日,3906 - 3918。doi: 10.1096 / fj.13 - 247635
费舍尔,T。Hayn,。,和Mierke, C. T. (2020). Effect of nuclear stiffness on cell mechanics and migration of human breast cancer cells.前面。细胞Dev。杂志。8,393。doi: 10.3389 / fcell.2020.00393
霜,B。,Bardai, F. H., and Feany, M. B. (2016). Lamin dysfunction mediates neurodegeneration in tauopathies.咕咕叫。医学杂志。26日,129 - 136。doi: 10.1016 / j.cub.2015.11.039
霜,B。,Hemberg, M., Lewis, J., and Feany, M. B. (2014). Tau promotes neurodegeneration through global chromatin relaxation.Nat。>。17日,357 - 366。doi: 10.1038 / nn.3639
Fulga, t。,Elson-Schwab, I., Khurana, V., Steinhilb, M. L., Spires, T. L., Hyman, B. T., et al. (2007). Abnormal bundling and accumulation of F-actin mediates tau-induced neuronal degeneration在活的有机体内。Nat,细胞生物。9日,139 - 148。doi: 10.1038 / ncb1528
Gaub, b . M。,Kasuba, K. C., Mace, E., Strittmatter, T., Laskowski, P. R., Geissler, S. A., et al. (2020). Neurons differentiate magnitude and location of mechanical stimuli.Proc。国家的。学会科学。117年,848 - 856。doi: 10.1073 / pnas.1909933117
吉尔伯特·h·t·J。Mallikarjun, V。Dobre, O。,Jackson, M. R., Pedley, R., Gilmore, A. P., et al. (2019). Nuclear decoupling is part of a rapid protein-level cellular response to high-intensity mechanical loading.Commun Nat。10日,4149年。doi: 10.1038 / s41467 - 019 - 11923 - 1
Grashoff C。,Hoffman, B. D., Brenner, M. D., Zhou, R., Parsons, M., Yang, M. T., et al. (2010). Measuring mechanical tension across vinculin reveals regulation of focal adhesion dynamics.自然466年,263 - 266。doi: 10.1038 / nature09198
Guilluy C。,Osborne, L. D., van Landeghem, L., Sharek, L., Superfine, R., Garcia-Mata, R., et al. (2014). Isolated nuclei adapt to force and reveal a mechanotransduction pathway in the nucleus.Nat,细胞生物。16 (4),376 - 381。doi: 10.1038 / ncb2927
Hamouda, m . S。,Labouesse, C., and Chalut, K. J. (2020). Nuclear mechanotransduction in stem cells.咕咕叫。当今。细胞生物。64年,97 - 104。doi: 10.1016 / j.ceb.2020.05.005
他,h·J。,Wang, X. S., Pan, R., Wang, D. L., Liu, M. N., and He, R. Q. (2009). The proline-rich domain of tau plays a role in interactions with actin.BMC细胞杂志。10日,81年。doi: 10.1186 / 1471-2121-10-81
Hervieu g J。,Cluderay, J. E., Gray, C. W., Green, P. J., Ranson, J. L., Randall, A. D., et al. (2001). Distribution and expression of TREK-1, a two-pore-domain potassium channel, in the adult rat CNS.神经科学103年,899 - 919。doi: 10.1016 / s0306 - 4522 (01) 00030 - 6
胡,X。,Liu, Z. Z., Chen, X., Schulz, V. P., Kumar, A., Hartman, A. A., et al. (2019). MKL1-actin pathway restricts chromatin accessibility and prevents mature pluripotency activation.Commun Nat。10日,1695年。doi: 10.1038 / s41467 - 019 - 09636 - 6
赫顿,M。,Lendon, C. L., Rizzu, P., Baker, M., Froelich, S., Houlden, H., et al. (1998). Association of missense and 5′-splice-site mutations in tau with the inherited dementia FTDP-17.自然393年,702 - 705。doi: 10.1038/31508
Janota c。,Calero-Cuenca, F. J., and Gomes, E. R. (2020). The role of the cell nucleus in mechanotransduction.咕咕叫。当今。细胞生物。63年,204 - 211。doi: 10.1016 / j.ceb.2020.03.001
贾,Y。,Vong, J. S.-L., Asafova, A., Garvalov, B. K., Caputo, L., Cordero, J., et al. (2019). Lamin B1 loss promotes lung cancer development and metastasis by epigenetic derepression of RET.j . Exp。地中海。216年,1377 - 1395。doi: 10.1084 / jem.20181394
江,L。,和Wolozin, B. (2021). Oligomeric tau disrupts nuclear envelope via binding to lamin proteins and lamin B receptor.阿尔茨海默氏痴呆17日,e054521。doi: 10.1002 / alz.054521
科比,t·J。,和Lammerding, J。(2018). Emerging views of the nucleus as a cellular mechanosensor.Nat,细胞生物。20岁,373 - 381。doi: 10.1038 / s41556 - 018 - 0038 - y
栗山,S。,Theveneau, E., Benedetto, A., Parsons, M., Tanaka, M., Charras, G., et al. (2014).在活的有机体内集体组织细胞迁移需要一个LPAR2-dependent增加流动性。j .细胞杂志。206年,113 - 127。doi: 10.1083 / jcb.201402093
Lammerding, J。方,l·G。霁,j . Y。Reue, K。,Stewart, C. L., Young, S. G., et al. (2006). Lamins A and C but not lamin B1 regulate nuclear mechanics.生物。化学。281年,25768 - 25780。doi: 10.1074 / jbc.M513511200
Lammerding, J。,Schulze, P. C., Takahashi, T., Kozlov, S., Sullivan, T., Kamm, R. D., et al. (2004). Lamin A/C deficiency causes defective nuclear mechanics and mechanotransduction.j .中国。调查113年,370 - 378。doi: 10.1172 / JCI19670
马奥尼,R。,Ochoa Thomas, E., Ramirez, P., Miller, H. E., Beckmann, A., Zuniga, G., et al. (2020). Pathogenic tau causes a toxic depletion of nuclear calcium.细胞的代表。32岁,107900年。doi: 10.1016 / j.celrep.2020.107900
Maingret F。Fosset, M。勒,F。,Lazdunski, M., and Honoré, E. (1999). TRAAK is a mammalian neuronal mechano-gated K+Channel.生物。化学。274年,1381 - 1387。doi: 10.1074 / jbc.274.3.1381
毛雷尔,M。,和Lammerding, J。(2019). The driving force: Nuclear mechanotransduction in cellular function, fate, and disease.为基础。启生物医学。Eng。21日,443 - 468。doi: 10.1146 / annurev - bioeng - 060418 - 052139
Montalbano, M。麦卡伦,S。,Sengupta, U., Puangmalai, N., Bhatt, N., Ellsworth, A., et al. (2019). Tau oligomers mediate aggregation of RNA-binding proteins Musashi1 and Musashi2 inducing Lamin alteration.衰老细胞18日,e13035。doi: 10.1111 / acel.13035
纳瓦,M . M。,Miroshnikova, Y. A., Biggs, L. C., Whitefield, D. B., Metge, F., Boucas, J., et al. (2020). Heterochromatin-driven nuclear softening protects the genome against mechanical stress-induced damage.细胞181年,800 - 817。e22。doi: 10.1016 / j.cell.2020.03.052
Ngai, a . C。,和Winn, H. R. (1996). Estimation of shear and flow rates in pial arterioles during somatosensory stimulation.点。j . Physiology-Heart循环生理270年,H1712-H1717。doi: 10.1152 / ajpheart.1996.270.5.H1712
Padmakumar, v . C。亚伯拉罕,S。、Braune年代。,Noegel, A. A., Tunggal, B., Karakesisoglou, I., et al. (2004). Enaptin, a giant actin-binding protein, is an element of the nuclear membrane and the actin cytoskeleton.Exp。细胞Res。295年,330 - 339。doi: 10.1016 / j.yexcr.2004.01.014
Paonessa F。,Evans, L. D., Solanki, R., Larrieu, D., Wray, S., Hardy, J., et al. (2019). Microtubules deform the nuclear membrane and disrupt nucleocytoplasmic transport in tau-mediated frontotemporal dementia.细胞的代表。26日,582 - 593。e5。doi: 10.1016 / j.celrep.2018.12.085
Pothapragada, s P。古普塔,P。,Mukherjee, S., and Das, T. (2022). Matrix mechanics regulates epithelial defence against cancer by tuning dynamic localization of filamin.Commun Nat。13日,218年。doi: 10.1038 / s41467 - 021 - 27896 - z
Prissette, M。,愤怒,W。高斯,M。,Racioppi, C., Fedorova, D., Dragileva, E., et al. (2022). Disruption of nuclear envelope integrity as a possible initiating event in tauopathies.细胞的代表。40,111249年。doi: 10.1016 / j.celrep.2022.111249
斯佩克特,我。,Shochet, N. R., Kashman, Y., and Groweiss, A. (1983). Latrunculins: Novel marine toxins that disrupt microfilament organization in cultured cells.科学219年,493 - 495。doi: 10.1126 / science.6681676
Steinhilb, m . L。,Dias-Santagata, D., Mulkearns, E. E., Shulman, J. M., Biernat, J., Mandelkow, E. M., et al. (2007). S/P and T/P phosphorylation is critical for tau neurotoxicity in Drosophila.j . >。Res。85年,1271 - 1278。doi: 10.1002 / JNR.21232
沃克,c . J。Crocini C。拉米雷斯,D。,Killaars, A. R., Grim, J. C., Aguado, B. A., et al. (2021). Nuclear mechanosensing drives chromatin remodelling in persistently activated fibroblasts.Nat,生物医学。Eng。5,1485 - 1499。doi: 10.1038 / s41551 - 021 - 00709 - w
维c W。,Wszolek, M. F., Shulman, J. M., Salvaterra, P. M., Lewis, J., Hutton, M., et al. (2001). Tauopathy in Drosophila: Neurodegeneration without neurofibrillary tangles.科学293年,711 - 714。doi: 10.1126 / SCIENCE.1062382
关键词:tauopathy、烦恼、核紧张,阿尔茨海默病,细胞核,力学生物学
引用:孙C,马J,射线WJ和霜B(2023)致病τ减少核紧张局势培养神经元。前面。老化4:1058968。doi: 10.3389 / fragi.2023.1058968
收到:2022年9月30日;接受:2023年1月11日;
发表:2023年1月23日。
编辑:
内森•伍德尔英国格拉斯哥大学版权孙©2023,妈,雷和霜冻。这是一个开放分布式根据文章知识共享归属许可(CC)。使用、分发或复制在其他论坛是允许的,提供了原始作者(年代)和著作权人(s)认为,最初发表在这个期刊引用,按照公认的学术实践。没有使用、分发或复制是不符合这些条件的允许。
*通信:贝丝霜,bfrost@uthscsa.edu