竞争力和共生效率的紫花苜蓿根瘤菌favelukesiiORY1菌株的同源基因肽酶HrrP被确定和SapA负面影响的共生关系
 Andres Berais-Rubio
1 __,
Andres Berais-Rubio
1 __, 玛丽亚·a·莫雷尔Revetria
1、2、3,
玛丽亚·a·莫雷尔Revetria
1、2、3, Matias Gimenez
3、4中,
Matias Gimenez
3、4中, 圣地亚哥Signorelli
1、5 __和
圣地亚哥Signorelli
1、5 __和 豪尔赫·蒙扎
1 * __
豪尔赫·蒙扎
1 * __
- 1Laboratorio de Bioquimica Facultad de Agronomia大学de la那时,乌拉圭的蒙得维的亚
- 2Laboratorio de Microbiologia de Suelos Facultad de Ciencias大学de la那时,乌拉圭的蒙得维的亚
- 3Laboratorio Microbiologia分子、Departamento Bioquimica y Genomica Microbianas (BIOGEM),学院Investigaciones Biologicas克莱门特Estable (IIBCE),乌拉圭蒙得维的亚
- 4Laboratorio de Genomica Microbiana,巴斯德研究所,乌拉圭蒙得维的亚
- 5能源植物生物学、分子科学学院、西澳大利亚大学,克劳利,佤邦,澳大利亚
在酸性土壤,根瘤菌favelukesii菌株,称为Oregon-like菌株,是苜蓿生产的潜在风险考虑到他们寄生行为。在这项研究中,我们分离五寄生菌株(ORY1 ORY5)从紫花苜蓿根瘤生长在乌拉圭酸性土壤,以99.7%和100%的16 s rRNA基因序列的身份r . favelukesiiLUP83的模式菌株。五项隔离箱资料显示两种不同的模式,表明一些耐酸多样性之间的隔离。基因组序列分析r . favelukesii菌株ORY1、LPU83 Or191表明他们有大约87.5%的常见的编码基因,包括共生基因。此外,ORY1共生基因的系统发育分析nifH,nifD,nifK,野田佳彦,nodB,诺德的共生基因有关吗大肠meliloti。我们证人ORY1竞争力通过接种种子99:1 ORY1和1:9 9比率::gusA/大肠melilotiU143。在这两种治疗方法,ORY1::gusA占据超过50%的结节,证明其高竞争力。然而,这些疗法的空中生物量显著不同,这表明有效应变引起的结节是必要的为最优植物生长提供足够的N。这些发现支持需要接种效率低下压力的地区可能存在。最后,我们发现三个基因编码的氨基酸序列域M16肽酶(同源性细菌hrrP和此类的基因),其中两个相邻,位于一个辅助质粒,而另一个是一个染色体基因。这些基因可能参与ORY1应变的寄生行为。
介绍
失败在紫花苜蓿(紫花苜蓿)生产归因于土壤酸性pH值、铝毒性(小结,2012;贾斯瓦尔et al ., 2018;施et al ., 2022)和寄生虫的存在效率低下结瘤苜蓿根瘤菌菌株(托雷斯Tejerizo et al ., 2011;赶et al ., 2022)。这些寄生虫菌株,称为Oregon-like,耐酸性的条件下,宿主范围广泛(赶et al ., 1992;Del Papa et al ., 1999),它的存在也被证实在美国和加拿大(布罗姆菲尔德et al ., 2010)。同样,俄勒冈州应变LPU83已经从紫花苜蓿根孤立结节在阿根廷和被分配到小说的物种根瘤菌favelukesii(托雷斯Tejerizo et al ., 2016)。俄勒冈州的菌株有竞争力的有节紫花苜蓿在酸性土壤和在这豆科植物共生效率低(Del Papa et al ., 2003;托雷斯Tejerizo et al ., 2011;赶et al ., 2022)。出于这个原因,重要的是要知道存在这种类型的应变在紫花苜蓿种植的地方,以及他们的竞争力对变质剂使用或本地高效菌株在土壤。
此外,下属机制解释缺乏固氮的寄生虫或低效品种基本上仍是无人。然而,在Ensifer meliloti medicago truncatula共生,大肠meliloti低效品种,低效率的机制是实现解释为根瘤菌的肽酶负面影响共生通信(价格et al ., 2015)。特别是,骗子et al。(2012)证明了效率低下大肠meliloti菌株能够获得兼容性时失去了宿主范围(人力资源)辅助质粒。反过来,高效菌株获得了人力资源质粒时变得效率低下。大约10%的Ensifersp.隔离估计进行辅助质粒基因,命名宿主范围限制肽酶(hrrP),编码的M16 metallopeptidase水解植物产生结节丰富的半胱氨酸肽(NCR)。在m . truncatula发现,约600名NCR肽(周et al ., 2013)。其中一个肽,NCR169,已经被证明是必要的类细菌分化的结节。m . truncatulaKO-mutants NCR169肽,dnf7,无法引起类细菌发展,这表明NCR169肽发挥作用的共生通信(霍et al ., 2015)。同样的,此类的(symbiotic-associated肽酶)chromosomic基因的大肠meliloti被证明编码HrrP-like M16肽酶,以类似的方式调节共生(本尼迪克特et al ., 2021)。在这里,我们报告中这些基因的存在Oregon-type紧张,这可能导致他们可怜的理解共生效率。
在乌拉圭土壤、竞争和无效的结瘤苜蓿根瘤菌菌株被发现并建议Oregon-like应变,但没有证实(Castro-Sowinsky et al ., 2002)。这些寄生虫苜蓿植入菌株构成潜在风险(尼尔森et al ., 2019),即使他们是共存与高效的共生体大肠meliloti。因此,在这项研究中,我们为苜蓿低效的根瘤菌菌株分离植物生长在酸性土壤,旨在证实在乌拉圭Oregon-like菌株的存在;评估这些菌株的竞争力;和M16-like肽酶识别的机制可以解释这种寄生虫的行为。
材料和方法
细菌和生长条件
耐酸(pH值5.2)根瘤菌菌株孤立结节的紫花苜蓿生长在酸性土壤在乌拉圭(表1),获得生物化学实验室的根瘤菌收集农学的学校,大学de la那时,乌拉圭。Or191来自俄勒冈州,美国(赶,大卫,1985从阿根廷)和LPU83 (托雷斯Tejerizo et al ., 2016)被用作参考的根瘤菌favelukesii。Ensifer melilotiU143 (= MCH3)应变,在乌拉圭苜蓿接种菌(Altier et al ., 2013)是由微生物实验室(Ministerio de Ganaderia水资源y Pesca)。根瘤菌和大肠meliloti是生长在YEM 28°C (文森特,1970)。大肠杆菌应变S17-1ʎpir含有质粒与转座子pCAM131 mTn5SSgusA31 (威尔逊et al ., 1995)生长在37°C磅(米勒,1972)补充壮观霉素(Sp) 100 ug /毫升和链霉素(St) 50µg /毫升。在液体媒体120 rpm的文化都摇动了。根瘤菌和大肠杆菌菌株保存在4°C YEM琼脂或磅琼脂Sp 100µg /毫升,分别。

表1位置和土壤的pH值m .漂白亚麻纤维卷低效的根瘤菌菌株被孤立。
植物和生长条件
紫花苜蓿简历。长安汽车种子表面消毒根据(Irisarri et al ., 2019)。植物生长状况控制在23/20°C(白天/晚上),16/8 h光周期和220µE.m2。s1光合光量子通量密度。
执行时间有节试验在管20毫升Jensen介质(詹森,1942摘要。竞争力,共生效率和相对共生效益分析进行在350毫升与无菌蛭石-砂锅(1:1),浇水或者蒸馏水和Fahraeus介质(文森特,1970)。蒸馏水和Fahraeus介质pH值调整到5.5 MES 10毫米。接种在每个试验进行了如下所述。
标签根瘤菌ORY1应变gusA基因
进行了结合使用根瘤菌菌株ORY1作为受体大肠杆菌应变S17-1λ-pir包含pCAM131,转座子mTn5SSgusA31作为供体,通常使用的过程Irisarri et al。(2019)。混合物保持在28°C隔夜与泰板(贝灵哲酒庄,1974)。Transconjugants选择板YEM Sp 100µg /毫升和呋喃妥英(Ntf) 20µg /毫升抑制大肠杆菌增长。
Transconjugants时间有节化验
时间有节化验三transconjugants尊重父母的压力是评价根据出现的第一个结节,结节在21天。pre-germinated种子,播种率每管Jensen介质和8个重复治疗,注射100µL细菌悬液的OD620年= 0.9 (108生/毫升)。ORY1::gusA克隆选择的竞争分析。
格斯活动苜蓿结节
结节状的根用蒸馏水洗净,与0.1磷酸盐缓冲剂pH = 7,孵化16 h在37°C在黑暗中包含1% SDS溶液中的类型,EDTA pH值0.5米8和1毫米5-bromo-4-cloro-3indolil-β-D-glucuronide (X-gluc)在50 mM磷酸钠缓冲pH值7.5 (威尔逊et al ., 1995)。结节被ORY1::gusA被蓝染色,视觉识别与那些被non-tagged菌株感染仍是清白的(Irisarri et al ., 2019)。
竞争力和生物质生产试验
竞争力U143和ORY1::gusA菌株进行了与sand-vermiculite锅(1:1),5植物/锅,4复制/治疗。三化验,n = 4和12个结果复制池进行了统计分析。锅是用水冲洗或Fahraeus介质pH值5.5,如上所示。菌株生长在YEM介质OD620年= 0.9。竞争力的实验包括ORY1::gusA和U143 99:1和1:9 9比率(名为治疗分别为99%和1%)。细菌浓度是根据确定的Riviezzi et al。(2020)。植物收获后21天接种和洗根结节染色如上所述。评估共生效率、生物质生产决定拍摄干重60°C 72 h,包括两个控件:U143应变和未经变质处理的治疗。
相对共生的有效性分析
种子被播种的速度5每锅和接种ORY1 U143菌株如前所述,和收获接种后35天。每个治疗包括5复制。相对共生效率(%)是基于ORY1寄生应变射干重生产相对于有效U143应变。
基因组DNA提取
从不同rhizobial菌株基因组DNA提取提取DNA提取设备(试剂盒,Alemania)。DNA的质量和浓度测定Nanodrop 2000(美国热科学),其完整性是可视化在琼脂糖凝胶1.2%。
16 srna基因扩增
16 s rRNA部分基因放大使用引物27 f和1525 r和所描述的条件巷(1991)。PCR产品运行在100 V Tris-acetate缓冲pH值8.2和可视化在1.2%琼脂糖凝胶染色SYBR安全DNA凝胶染色(美国热费希尔科学)。Accuruler 1 kb DNA梯(Maestrogen公司、台湾)作为分子量标记。测序进行Macrogen Inc .(韩国)。
盒基因指纹
总基因组DNA扩增BOXA1底漆(Koeuth et al ., 1995)。放大条件是1的循环使用2分钟在95°C, 30周期45赛格在93°C, 1分钟50°C和6分钟在65°C,和最后一个扩展的8分钟68°C。PCR产品可视化如前所述。
基因组测序
基因组测序获得使用Illumina公司技术(Macrogen、韩国)收益率1110万paired-end读取长度为151个基点。低质量的读取与参数使用Trimmomatic软件过滤掉2:30:10领先:3落后:3 SLIDINGWINDOW: 15 MINLEN: 75 (博尔格et al ., 2014)。软件是用来执行一个黑桃新创装配使用剩下的配对阅读(Bankevich et al ., 2012)。组装的质量评估与QUAST软件使用LPU83基因组作为引用(维奇et al ., 2013)。质粒重叠群被发现使用plaSquid软件(Gimenez et al ., 2022)选项-minidist和使用LPU83质粒作为参考数据库。基因组注释的所有分析基因组与默认参数(使用prokka软件完成Seemann 2014)。
计算的平均核苷酸指数(ANI),相关索引tetra-nucleotide签名和利乐相关搜索(TCS)进行使用JSpeciesWSserver (里希特et al ., 2016)。基因组序列数据上传到基因组服务器类型(双柄陶制大酒杯),整整一个genome-based分类分析(Meier-Kolthoff完全懂得,2019)。
系统发育分析
共生基因nifH、nifD nifK,野田佳彦,nodB和诺德,从ORY1基因组序列提取。获得的序列与参考株序列可用在公共场合国家生物技术信息中心(NCBI)的数据库。序列与ClustalW算法和最大似然phylogenomic树(田村和Nei, 1993年每个茎)构造序列分析(二)。比对和树木建造了大型11软件(田村et al ., 2021)。统计支持树节点被引导分析评估使用1000复制(Felsenstein, 1985)。
加入核苷酸序列号码
16 s rRNA的基因库加入数字序列用于系统发育分析:根瘤菌favelukesiiORY1 (OP294988) ORY2 (OP294989) ORY3 (OP294990) ORY4 (OP294991) ORY5 (OP294992) Or191 (EU928874.1),根瘤菌anhuiensePVPR1 (MT476932.1),根瘤菌indigoferaeNAC94 (MK872365.1),Bradyrhizobium yuanmingenseVAFW14 (LC585439.1),根瘤菌croatiense9 t (MK753104.1),根瘤菌altiplani152084.1 BR 10423 (NR),根瘤菌tibeticumCCBAU85039 (NR116254.1)和Sinorhizobium meliloti2011 (CP004140.1)。
加入基因库数字基因和质粒的共生基因和部分16 s序列提取在括号中表示:根瘤菌favelukesiiORY1 (JAIRAY000000000.1);LPU83 (HG916852.1)和Or191 (GCA_000419725.1);根瘤菌tibeticumCCBAU85039 (GCF_900108425.1);根瘤菌altiplani10423 BR (GCA001542405.1);根瘤菌grahamiiBG7 (CP043498.1);和Sinorhizobium meliloti2011 (CP004140.1);根瘤菌tropici国际热带农业中心899 020061.1 (NC);根瘤菌esperanzaeN561 (CP013501.1);根瘤菌phaseoliR650 (CP013533.1);根瘤菌gallicumsp。R602 (CP006878.1);根瘤菌etliNXC12 (CP020907.1)。
根瘤菌plasmidic肽酶分析
Ensifer melilotiUSDA1963肽酶HrrP从NCBI参考蛋白质数据库检索和用于检测其在web服务器(包含域贝特曼et al ., 2004)。隐马尔可夫模型从M16肽酶(PF00675)和M16_C肽酶(PF05193)包含了从数据库下载。理解的存在潜在的肽酶质粒从根瘤菌属,461个质粒从PLSDB下载数据库(v . 2021 _06_23_v2) (Galata et al ., 2019)。所有编码蛋白质利用prokka预测软件。HMMER 3.0软件被用来扫描M16肽酶域使用的选项-cut_ga作为过滤阈值(艾迪,2011)。预测蛋白质都聚集到98%的身份,使用CD-HIT软件,减少冗余序列(傅et al ., 2012)。r . favelukesii预测M16肽酶被添加到系统发育分析。序列(可用:https://doi.org/10.6084/m9.figshare.21641816.v1)使用肌肉对齐算法通过msa R包(Bodenhofer et al ., 2015)。系统发育树与猿使用neighbor-joining算法计算R包(- et al ., 2004)和用ggtree (Yu et al ., 2017)。域组织每个序列的手动从数据库搜索和添加到包含了分析。
统计分析
方差分析是在确定统计学意义和R软件事后测试(图基´s测试)被用于成对比较来确定P值。
结果
ORY1 alfalfa-parasite根瘤菌菌株根瘤菌favelukesii
来描述m .漂白亚麻纤维卷寄生虫根瘤菌ORY1、ORY2 ORY3、ORY4 ORY5菌株,从乌拉圭分离酸性土壤,我们分析了盒,16 s rRNA的序列,和相对共生效率。
箱资料五原地隔离和阿根廷和美国r . favelukesii菌株表明ORY1和ORY5菌株有相同的形象外交压力,而ORY2, ORY3,和ORY4菌株相同的概要文件,但是不同的外国压力(图1一个)。

图1多样性和系统发育的关系r . favelukesii菌株。(一)盒子的r . favelukesii菌株。ORY1、ORY2 ORY3, ORY4 ORY5收集菌株在乌拉圭酸土壤和参考菌株LUP83和Or191酸分别Argentinan和美国土壤。微波加工(分子量标记)。(B)种系发生树推断使用最大似然方法基于16 s rRNA基因的部分序列比对(1.342元)。用粗体字母表示压力从土壤乌拉圭孤立。其他菌株的基因组从NCBI获得。星号(*)表示应变类型。1000复制引导计算值。酒吧,每100元1元替换。
部分16 s rRNA ORY1基因序列,ORY4 ORY5 100%相同R。favelukesii菌株,LPU83 Or191,与阿根廷和美国分别酸性土壤。ORY2 ORY3菌株显示,分别有99.9%和99.7%的身份ORY1, LPU83, Or191。接下来,我们进行了系统发育分析不同菌株与乌拉圭的土壤(ORY)以及其他相关的物种。所有ORY菌株聚在一起R。favelukesiiLPU83 Or191菌株,密切相关r . tibeticum(图1 b)。此后,ORY1应变的应变作为研究进一步化验。的寄生行为证实了ORY1射干重生产这一毒株为唯一变质剂的存在。这个参数是只有15%的这样的获得商业接种菌菌株U143,证明共生效率非常低。
ORY1菌株的基因组测序和系统发育分析
草案的基因组分析r . favelukesiiORY1应变表明,基因组的大小是7.44 Mb;有59.7%的GC内容;包含7509个编码和51 RNA基因。五经由被确定对应染色体和四个质粒。质粒的分析显示,151年叠连群总长度的96%根瘤菌favelukesiiLPU83质粒。这代表了总长度3.25 Mb的orf编码3349。基于TCS或tetra-nucleotide签名,只有r . favelukesii和r . tibeticum被发现与z得分> 0.999密切相关。ANI值> 96%一致组菌株的基因组r . tibeticum和r . favelukesii加上ORY1,但识别ORY1 ANI值> 99%r . favelukesii。根据双柄陶制大酒杯ORY1属于分析r . favelukesii。的系统发育分析r . favelukesiiORY1、Or191 LPU83菌株显示,他们聚集在一起,分开r . tibeticum当使用完整的基因组(图2一个)。此外,r . favelukesii和r . tibeticum也将分开r . grahamii和r . altiplani(图2一个)。同样的,r . favelukesii菌株不组r . tibeticum当一个二的共生基因,nifK nifD nifH,野田佳彦nodB,诺德,被认为是。此外,这些连接相关的基因大肠meliloti共生基因(图2 b)。为了更好地理解这三个之间的相似r . favelukesii菌株,ORY1 Or191 LPU83,我们之间的重叠分析三个完整基因组通过识别同源基因。我们观察到,大多数编码基因是常见的所有菌株(87.5%左右,相对于ORY1) Or191有更大的重叠ORY1比LPU83 ORY1独特和大约2.7%的编码基因(图2 c)。

图2系统发育分析和同源基因在三维恩图r . favelukesii菌株。(一)从基因组树推断爆炸距离发展史(GBDP)距离计算从基因组序列与双柄陶制大酒杯(https://tygs.dsmz.de/)。分支长度比例GBDP距离。上面的数字分支GBDP pseudo-bootstrap支持从100年复制值> 60%,平均83.8%的分支的支持。这棵树扎根在中点。(B)二共生基因nifD、nifK nifH,野田佳彦nodB,诺德不同根瘤菌的物种。(C)在三维恩图的同源基因r . favelukesii菌株。重叠区域代表pan-genomes之间共同同源基因。
存在的竞争和共生效率ORY1高效U143压力
评估ORY1应变的竞争力我们标记ORY1报告基因,gusA。确认的gusA插入并不影响竞争力和生物质生产,我们比较ORY1的性能:gusA这些参数应变,ORY1父母的压力。没有观察到这些菌株之间的差异时间观察第一个结节(8天),每个工厂的结节数21天,生物质开枪。
两种治疗方法被用来测试竞争力和生物质生产,ORY1组成的混合和商业变质剂,大肠melilotiU143。一个治疗由99:1比(ORY1::gusA/ U143以下99%疟原虫治疗),而其他的治疗方法是1:9 9比率(ORY1::gusA/ U143,以下1%疟原虫治疗)。结节占领ORY1::gusA应变99%治疗大于1% (图3一)。值得注意的是,即使在治疗,1%的比例占据结节超过总额的50%的结节,证明这一毒株的竞争力。尽管相当类似的职业治疗,观察显著不同的生物质生产治疗(99%和1%之间图3 b)。事实上,生物质生产99%的治疗方法是非常低的,而不是统计不同获得的徵控制,而生物质生产的1%治疗没有统计学上不同的获得与U143应变(图3 b)。无论寄生虫占据的百分比结节,结节的总数是更大的治疗(99%图3 c)。尽管更多的结节的99%治疗,白色结节的数量也显著大于1%的治疗(图3 d)。
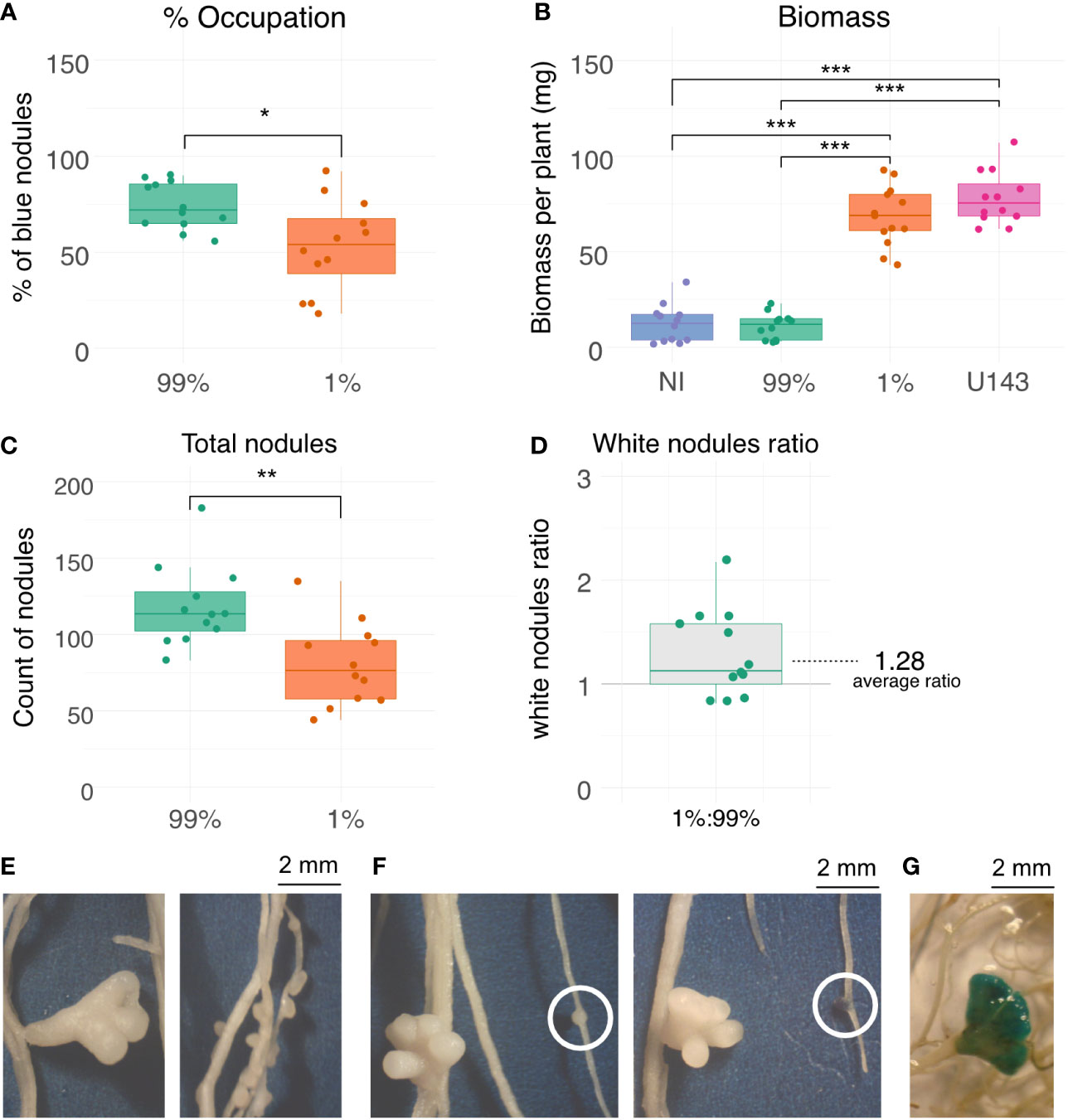
图3竞争力,生物质生产和结节的形态与寄生植物接种高效菌株。(一)结节占领标记菌株ORY1百分率:gusA对总结节。(B)空中生物质生产。(C)总结节的数量。(D)白色结节总比。(E)那些被低效和高效菌株的结节方面(左和右,分别)。(F)GUS染色的结节(左前染色,染色后)。(G)GUS染色的结节呈现不同的形态(U143诱导morphology-like)。99%和1%对应co-inoculations 99:1比例和ORY1 1:9 9::gusA和U143分别;倪徵控制;U143,接种有效应变作为商业变质剂。*,p值< 0.05;* *,p值< 0.01;* * *,p值< 0.001。
结节形态由ORY1::gusA和U143菌株不同(图3 e)。我们发现ORY1::gusA诱发大量的发展小球形结节与U143不同,导致少数较大的待定结节(图3 f)。异常,少数U143-occupied-like结节被ORY1::gusA(图3 g),这一现象我们属性co-occupation这些菌株,鉴于我们从来没有观察到这种形态当植物完全接种ORY1::gusA。
识别的M16肽酶域ORY1压力
考虑到细菌的基因hrrP和此类的编码的M16肽酶与根瘤菌共生效率较低,我们寻求基因编码M16肽酶在ORY1基因组中。生物信息学分析的蛋白质域使用HMMER软件,结合蛋白质数据库包含了允许我们证据ORY1基因组的存在三个基因编码aminoacidic序列M16锌metallopeptidase域。这些基因的两个相邻基因位于一个辅助质粒,和另一个是chromosomic基因。
我们NCBI蛋白质执行搜索,发现532 448 07295 (ID)和07296年(ID) aminoacidic连续的序列编码ORY1辅助质粒的菌株显示身份的26.55%和22.78,50 - 63%的覆盖率,分别的HrrP肽酶E。melilotiUSDA1963 (表2)。这种蛋白质是一种homodimeric M16锌metallopeptidase参与NCR肽的乳沟。染色体的ORY1应变,我们发现了一个DNA序列编码432个胺基酸序列呈现一个82.78%的身份和98%的覆盖率SapA肽酶大肠meliloti1021株(CAC45492.2)。这大肠melilotiSapA属于保守M16B亚科肽酶存在于许多不同的根瘤菌。

表2爆炸M16域分析中确定ORY1对参考序列。创造价值对应的数量预计达到类似的质量(分数),可以发现只是偶然。
在根瘤菌评估M16的基因的存在,我们分析了461个质粒可以在PLSDB数据库。只有30个基因编码这种蛋白领域被发现的,其中一些对应于相同的经由。M16 domain-encoding基因被发现在根瘤菌等不同种类的质粒r . leguminosarum,r . etli,r . favleukesii和r . gallicum(图4)。此外,集群的序列中发现这个系统发育树没有显示一个物种分布。

图4M16肽酶序列的系统发育树编码在质粒不同根瘤菌的物种。肽酶序列的系统发育树与Neighbourg-Joining计算算法。提示颜色表明根瘤菌宿主物种。尖的形状表明不同域架构,手动浏览每个肽酶序列包含了web服务器。
我们进一步了解远程同源性模式在这些基因分析领域架构的根瘤菌质粒编码的肽酶(图4)。我们观察到有不同的域组织但与有限数量的蛋白质域;specificallyM16 (PF00675)和M16_C (PF05193)。肽酶M16紧随其后肽酶M16_C域架构这些蛋白质,存在于大多数的至少一个基因5根瘤菌物种代表树的组织这一领域。无特征的参考蛋白质拥有此域名正在存入UniProt加入号码W2S8M8 AlphaFold结构预测。作为一个功能蛋白质我们包含了HrrP的特点E.meliloti美国农业部1963年,导致类似的领域体系结构只有一个额外的肽酶M16_C域。参考蛋白在这些领域也存入UniProt入世W9RIX3之下。
讨论
发展史和多样性的俄勒冈州乌拉圭酸土壤中分离到的
紫花苜蓿寄生Oregon-like根瘤菌是植入的问题和生产草原的豆类(托雷斯Tejerizo et al ., 2011)。这些根瘤菌被确认在美国、加拿大(赶et al ., 1985;布罗姆菲尔德et al ., 2010),在阿根廷,小说的物种r . favelukesii描述(托雷斯Tejerizo et al ., 2016)。Castro-Sowinsky et al。(2002)定意的存在Oregon-like根瘤菌在乌拉圭的土壤。然而,这根瘤菌在乌拉圭的存在土壤之前并没有得到证实。在这项研究中,基于五个分离株的全基因组测序分析来自乌拉圭酸性土壤,哪里有苜蓿生产产量问题,我们证实了俄勒冈州菌株的存在(图2一个)。
r . favelukesii和r . tibeticum可以结瘤m .漂白亚麻纤维卷(侯et al ., 2009),但r . tibeticum等多个主机吗生长archiducis-nicolai,Medicago lupulina,Phaseolous寻常的等(Abd-Alla et al ., 2014;托雷斯Tejerizo et al ., 2017)。在一致,我们的基因组为基础的系统发育分析表明,r . favelukesii菌株(ORY1 Or191和LPU83)聚集在一起但分开r . tibeticum应变CCBAU85039 (图2一个)。此外,基于共同的基因的基因组比较显示,ORY1应变比LPU83更类似于Or191孤立于美国与阿根廷(图2 c)。数量有限的独特基因ORY1(2.7%)大多是无特征基因和我们没有识别出任何共生基因。
我们的结果也显示更大的多样性比阿根廷酸性土壤中观察到。在我们的研究中,从乌拉圭酸性土壤中分离出五株,两个菌株呈现相同的盒子形象LPU83 Or191,在阿根廷和美国分别确定,而其他三个呈现不同的概要文件。与这个结果不同,十二从阿根廷酸性土壤分离导致同一ERIC指纹概要文件从美国Or191 (韦格纳et al ., 2001),代表一个非常低的遗传多样性。较低的遗传多样性是不同于通常为根瘤菌在地理上广泛分布的报道(Martinez-Romero Caballero-Mellado, 1996),我们观察clover-nodulating根瘤菌(Irisarri et al .,塔尔塔利亚et al ., 2019)和lotus-nodulating根瘤菌(Sotelo et al ., 2011;巴蒂斯塔et al ., 2013)与乌拉圭的土壤。尽管如此,十二个隔离测试韦格纳et al。(2001)选择从最初的465年阿根廷土壤隔绝,与pH值在5.5和6.2之间,因为生长在pH值5.0 (Del Papa et al ., 1999)。这意味着12孤立与相同的ERIC指纹被高容忍度低pH值选择,和极端环境条件减少遗传多样性,而乌拉圭土壤的pH值,只有我们的菌株略酸(pH值5.4 - 5.8),我们收集解释观察到的更大的遗传多样性。
紫花苜蓿co-inoculation ORY1 / U143:竞争力和生物质生产
高竞争力的寄生ORY1应变显示占用1%的结节治疗(图3一)。有趣的是,生物产量高,而不是不同的,U143 (图3 b),尽管超过50%的结节被寄生虫应变ORY1占领。这result suggests that the presence of parasite rhizobia should not be a problem for biomass production if our design (1:99, parasite:efficient rhizobia) represents the situation occurring during the first few days after sowing, where the strain used as commercial inoculants are expected to be at greater concentration than native parasite rhizobia. On the other hand, our results show that if the load of commercial inoculant is very low, or if the load of parasite rhizobia in the soil is high, severe problems in biomass production can be found (图3 b)。
乍一看,这听起来有悖常理,我们观察一个高生物质产量占领的疟原虫,ORY1,大于50%。这可以部分解释为1%治疗大约30%以上的白色(有效)结节治疗相对于99% (图3 d)。值得注意的是,不兼容的plasmid-carrying应变可以诱导的结节数量高于有效菌株,即使结果结节不太有利于主机(骗子et al ., 2012)。这是符合我们的观察,我们发现更多的总结节治疗丰富ORY1相对于1% (图3 c)。然而,结节寄生虫产生的应变被证明更有利于有效菌株产生的植物相比,这是建议产生更大更密集的结节(骗子et al ., 2012)。因此,如果我们只考虑白色根瘤固氮活性结节,这是不足为奇的1%治疗生物质生产大于99%的治疗。另一个因素可能占高生物量生产即使占领ORY1相对较高,与高效菌株co-occupation的可能性。Co-occupation一直在报道m . e . meliloti共生(马唐Checcucci et al ., 2016)。因此,我们假设许多蓝色结节也可以占领有效的应变导致固氮。事实上,我们观察到一些成熟的结节是疟原虫(图3 g),强烈建议这些结节co-occupation的可能性。因此,我们的观察评论的重要性不仅评估竞争力作为占领结节的比例,通常测试(Irisarri et al ., 2019),但也总而言,成熟的,不发达,co-occupied结节,更好地理解植物层次的结果。
识别假定的hrrP和此类的基因ORY1应变
的质粒中发现ORY1基因组编码的传染性假定的M16肽酶,在这三个高度保守的可用r . favelukesii的基因组。假定的M16肽酶的系统发育分析一组全面的根瘤菌质粒表明,r . favelukesii putatives肽酶不集群物种的系统发育树,类似于发生什么Ensifer序列(图4)。这提出了由于横向转移事件塑造这些质粒的进化历程肽酶(价格et al ., 2015)。
M16质粒肽酶在大肠meliloti和先前研究可以与主机限制函数(价格et al ., 2015)。这两个M16肽酶编码在r . favelukesii质粒是连续的和他们两人一起模拟域架构的大肠meliloti HrrP蛋白质,其中一个序列编码M16 - M16C域,和下一个序列M16C域。鉴于M16的邻近peptidase-encoding基因我们建议他们可能与hrrP作为一个基因融合或分裂事件的一部分,因为它已经在一组g +和革兰氏细菌(Pasek et al ., 2006)。此外,这些蛋白质可以隐含在类似的主机制约机制在大肠meliloti前面描述的物种。特别是HrrP和SapA肽酶发挥作用的消化NCR肽由主机,例如,似细菌的分化NCR169不可或缺的肽(霍et al ., 2015;价格et al ., 2015;本尼迪克特et al ., 2021)。这些肽所需的正确建立共生关系大肠meliloti - m . truncatula(价格et al ., 2015;本尼迪克特et al ., 2021)。的plasmidic肽酶HrrP降解NCR肽,将根瘤菌积极机制来控制主机(锅和王出版社,2017年;本尼迪克特et al ., 2021),其表达式通常与寄生虫有关的行为(价格et al ., 2015)。假定的存在hrrP和此类的基因在r . favelukesii俄勒冈州的菌株,ORY1 Or191, LPU83可以部分解释的寄生行为r . favelukesii证明了不寻常的球状的结节,hyper-nodulation和共生效率低。
结论
在乌拉圭,紫花苜蓿主要是播种在酸性土壤中,我们证实了俄勒冈州的存在寄生型菌株,盒子的资料让我们假设这个菌株的多样性高于之前报道。Co-inoculation化验与低寄生虫/高效菌株比率证明,一方面,ORY1应变比U143应变明显更有竞争力,另一方面,在第一阶段幼苗的生物量损失不是有意义的发展。这些发现支持的需要接种效率低下压力的地区可能存在。最后,我们描述了第一次hrrP的存在和sapA r . favelukesii基因,这可以部分解释俄勒冈州的寄生行为——比如菌株表现为结节不同寻常的球状的一面,hyper-nodulation和共生效率低。
数据可用性声明
在这项研究中提出的数据集可以在网上找到存储库。库的名称/存储库和加入号码可以找到(s)如下:https://www.ncbi.nlm.nih.gov/,JAIRAY000000000.1。
作者的贡献
作者证实对论文的贡献如下:研究概念和设计:JM。数据收集:AB-R, MG和毫米。分析和解释的结果:SS, AB-R, MG和MM, JM。草案手稿准备:AB-R,党卫军,JM。所有作者回顾了结果,批准了最终版本的手稿。所有作者的文章和批准提交的版本。
资金
作者感谢赠款的资助INIA L1四面八方4。PEDECIBA, Udelar-CAP (a . Berais-Rubio高地”格兰特)和SNI乌拉圭。
的利益冲突
作者声明,这项研究是在没有进行任何商业或财务关系可能被视为一个潜在的利益冲突。
出版商的注意
本文表达的所有索赔仅代表作者,不一定代表的附属组织,或出版商、编辑和审稿人。任何产品,可以评估在这篇文章中,或声称,可能是由其制造商,不保证或认可的出版商。
引用
Abd-Alla m . H。Issa A。,Ohyama T. (2014). “Impact of harsh environmental conditions on nodule formation and dinitrogen fixation of legumes,” in固氮作用的生物学和生态学的发展(伦敦:IntechOpen)卷。9。1。
Altier N。,Beyhaut E., Pérez C. (2013). “Root nodule and rhizosphere bacteria for forage legume growth promotion and disease management,” in细菌在土壤生物学:作物生产力(柏林,海德堡:Springer), 167 - 184。
Bankevich。Nurk S。,Antipov D., Gurevich A. A., Dvorkin M., Kulikov A. S., et al. (2012). SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing.j .第一版。医学杂志。19 (5),455 - 477。doi: 10.1089 / cmb.2012.0021
贝特曼。,硬币L。,Durbin R., Finn R. D., Hollich V., Griffiths-Jones S., et al. (2004). The pfam protein families database.核酸Res。32 (suppl_1), D138-D141。doi: 10.1093 / nar / gkh121
巴蒂斯塔L。,Tomasco I., Lorite M. J., Sanjuán J., Monza J. (2013). Diversity and phylogeny of rhizobial strains isolated fromLotus uliginosus生长在乌拉圭的土壤。达成。土壤。生态。66年,19-28。doi: 10.1016 / j.apsoil.2013.01.009
本尼迪克特a B。,Ghosh P。,Scott S. M., Griffitts J. S. (2021). A conserved rhizobial peptidase that interacts with host-derived symbiotic peptides.科学。代表。11 (1),1 - 10。doi: 10.1038 / s41598 - 021 - 91394 - x
贝灵哲酒庄j . e . (1974)。R因子转移根瘤菌leguminosarum。Microbiol将军。84年,188 - 198。doi: 10.1099 / 00221287-84-1-188
Bodenhofer U。,Bonatesta E., Horejš-Kainrath C., Hochreiter S. (2015). Msa: An r package for multiple sequence alignment.生物信息学31日(24),3997 - 3999。doi: 10.1093 /生物信息学/ btv494
博尔格a . M。Lohse M。,Usadel B. (2014). Trimmomatic: A flexible trimmer for illumina sequence data.生物信息学30 (15),2114 - 2120。doi: 10.1093 /生物信息学/ btu170
布罗姆菲尔德·e·s P。Tambong j . T。克劳蒂尔在S。,拉盖尔G。普雷沃斯特D。,Laguerre G., et al. (2010).Ensifer,Phyllobacterium和根瘤菌物种占领的结节紫花苜蓿(紫花苜蓿)和Melilotus阿尔巴(草木樨)生长在加拿大网站没有种植的历史。微生物学。156 (pt.2), 505 - 520。doi: 10.1099 / mic.0.034058-0
Castro-Sowinsky年代。,卡雷拉我。,Catalan A. I., Coll J., Martínez-Drets G. (2002). Occurrence, diversity and effectiveness of mid-acid tolerant alfalfa nodulating rhizobia in Uruguay.共生32岁,105 - 118。
Checcucci。,Azzarello E., Bazzicalupo M., Galardini M., Lagomarsino A., Mancuso S., et al. (2016). Mixed nodule infection inSinorhizobium meliloti- - - - - -紫花苜蓿共生建议的存在作弊行为。前面。植物科学。7日,835年。doi: 10.3389 / fpls.2016.00835
骗子m B。,Lindsay D. P., Biggs M. B., Bentley J. S., Price J. C., Clement S. C., et al. (2012). Rhizobial plasmids that cause impaired symbiotic nitrogen fixation and enhanced host invasion.摩尔。Plant-Microbe交互。25 (8),1026 - 1033。doi: 10.1094 / mpmi - 02 - 12 - 0052 - r
Del Papa m F。,Balague L. J., Castro-Sowinski S. C., Wegener C., Segundo E., Abarca F. M., et al. (1999). Isolation and characterization of alfalfa-nodulating rhizobia present in acidic soils of central Argentina and Uruguay.达成。环绕。Microbiol。65 (4),1420 - 1427。doi: 10.1128 / aem.65.4.1420 - 1427.1999
Del Papa m F。,Pistorio M。,Balagué L. J., Draghi W. O., Wegener C., Perticari A., et al. (2003). A microcosm study on the influence of pH and the host-plant on the soil persistence of two alfalfa-nodulating rhizobia with different saprophytic and symbiotic characteristics.医学杂志。土壤肥力39 (2),112 - 116。doi: 10.1007 / s00374 - 003 - 0690 - 6
赶b D。,David B. (1985). Characterization of rhizobia from ineffective alfalfa nodules: Ability to nodulate bean plants [菜豆萨维(l)。]T。达成。环绕。Microbiol。50 (6),1422 - 1427。doi: 10.1128 / aem.50.6.1422 - 1427.1985
赶b D。,Osman W. A. M., Ardley J., Zandberg J., Gollagher M., van Berkum P., et al. (2022). The genome of the acid soil-adapted strain rhizobium favelukesii OR191 encodes determinants for effective symbiotic interaction with both an inverted repeat lacking clade and a phaseoloid legume host.前面。Microbiol。13。doi: 10.3389 / fmicb.2022.735911
赶b D。,Young J. P. W., Selander R. K. (1992). Phylogenetic position of rhizobium sp. strain Or191 a symbiont of both紫花苜蓿和菜豆基于部分的序列16 s rRNA和nifH基因。达成。环境。Microbiol。58 (6),1809 - 1815。doi: 10.1128 / aem.58.6.1809 - 1815.1992
Felsenstein j . (1985)。信心限制的发展史:使用引导的方法。进化39 (4),783 - 791。doi: 10.1111 / j.1558-5646.1985.tb00420.x
傅L。,Niu B., Zhu Z., Wu S., Li W. (2012). CD-HIT: Accelerated for clustering the next-generation sequencing data.生物信息学28日,3150 - 3152。doi: 10.1093 /生物信息学/ bts565
Galata V。,Fehlmann T., Backes C., Keller A. (2019). PLSDB: A resource of complete bacterial plasmids.核酸Res。47 (D1) D195-D202。doi: 10.1093 / nar / gky1050
Gimenez M。,Ferrés I., Iraola G. (2022). Improved detection and classification of plasmids from circularized and fragmented assemblies.bioRxiv。doi: 10.1101 / 2022.08.04.502827
展开磋商。,Saveliev V., Vyahhi N., Tesler G. (2013). QUAST: Quality assessment tool for genome assemblies.生物信息学29 (8),1072 - 1075。doi: 10.1093 /生物信息学/ btt086
Horvath) B。,Domonkosa Á., Keresztb A., Szucs A., Ábrahámb E., Ayaydinc F., et al. (2015). Loss of the nodule-specific cysteine rich peptide, NCR169 abolishes symbiotic nitrogen fixation in the medicago truncatula dnf7 mutant.Proc。国家的。学会科学。美国112年,15232 - 15237。doi: 10.1073 / pnas.1500777112
侯b . C。,Wang E. T., Li Y. Jr., Jia R. Z., Chen W. F., Gao Y., et al. (2009).根瘤菌tibeticumsp. 11月,共生细菌隔绝生长archiducis- - - - - -尼科莱(Širj) vassilcz。进化Microbiol Int。j .系统性。59 (12),3051 - 3057。doi: 10.1099 / ijs.0.009647-0
Irisarri P。,Cardozo G., Tartaglia C., Reyno R., Gutiérrez P., Lattanzi F., et al. (2019). Selection of competitive and efficient rhizobia strains for white clover.前面。Microbiol。10。doi: 10.3389 / fmicb.2019.00768
贾斯瓦尔·K。,Naamala J。,Dakora F. D. (2018). Nature and mechanisms of aluminium toxicity, tolerance and amelioration in symbiotic legumes and rhizobia.医学杂志。Fertil。土壤54岁,309 - 318。doi: 10.1007 / s00374 - 018 - 1262 - 0
Koeuth T。,Versalovic J., Lupski J. R. (1995). Differential subsequence conservation of interspersed repetitive链球菌引起的肺炎框元素在不同的细菌。基因组Res。5 (4),408 - 418。doi: 10.1101 / gr.5.4.408.html
莱恩d . j . (1991)。“16 s / 23 s rRNA测序,”核酸技术在细菌分类学。Eds。Stackebrandt E。,Goodfellow M. (New York, NY: John Wiley and Sons).
Martinez-Romero E。,Caballero-Mellado J. (1996).根瘤菌的发展史和细菌遗传多样性。暴击。启植物科学。15 (2),113 - 140。doi: 10.1080 / 07352689.1996.10393183
Meier-Kolthoff j . P。,完全懂得m (2019)。双柄陶制大酒杯是一个自动化高通量平台先进的genome-based分类法。Commun Nat。10 (1),1 - 10。doi: 10.1038 / s41467 - 019 - 10210 - 3
尼尔森·j·F。,Castellani L. G., Draghi W. O., Pérez-Giménez J., Torres Tejerizo G. A., Pistorio M. (2019). Proteomic analysis of rhizobium favelukesii LPU83 in response to acid stress.j .蛋白质组Res。18 (10),3615 - 3629。doi: 10.1021 / acs.jproteome.9b00275
潘H。,Wang D. (2017). Nodule cysteine-rich peptides maintain a working balance during nitrogen-fixing symbiosis.Nat。植物3 (5),1 - 6。doi: 10.1038 / nplants.2017.48
- E。,Claude J., Strimmer K. (2004). APE: Analyses of phylogenetics and evolution inR语言。生物信息学20 (2),289 - 290。doi: 10.1093 /生物信息学/ btg412
Pasek年代。,Risler J. L., Brézellec P. (2006). Gene fusion/fission is a major contributor to evolution of multi-domain bacterial proteins.生物信息学22 (12),1418 - 1423。doi: 10.1093 /生物信息学/ btl135
价格p。,Tanner H. R., Dillon B. A., Shabab M., Walker G. C., Griffitts J. S. (2015). Rhizobial peptidase HrrP cleaves host-encoded signaling peptides and mediates symbiotic compatibility. Proc. Natl. Acad. Sci. U.S.A. 112, 15244–15249. doi: 10.1073/2Fpnas.1417797112
里希特米。,Rosselló-Móra R., Oliver Glöckner F., Peplies J. (2016). JSpeciesWS: A web server for prokaryotic species circumscription based on pairwise genome comparison.生物信息学32 (6),929 - 931。doi: 10.1093 /生物信息学/ btv681
Riviezzi B。,Cagide C。,Pereira A., Herrmann C., Lombide R., Lage M., et al. (2020). Improved nodulation and seed yield of soybean (Glycine max) with a new isoflavone-based inoculant ofBradyrhizobium elkanii。根际15日,100219年。doi: 10.1016 / j.rhisph.2020.100219
“H。,Sun G., Gou L., Guo Z. (2022). Rhizobia–legume symbiosis increases aluminum在紫花苜蓿耐。植物11(10),1275年。doi: 10.3390 / plants11101275
Sotelo M。,Irisarri P。,Lorite M., Casaretto E., Rebuffo M., Sanjuán J., et al. (2011). Diversity of rhizobia nodulatingLotus corniculatus生长在乌拉圭的北部和南部地区。达成。土壤生态。49岁,197 - 207。doi: 10.1016 / j.apsoil.2011.05.005
田村K。,Nei M. (1993). Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees.摩尔。杂志。另一个星球。10 (3),512 - 526。doi: 10.1093 / oxfo雷竞技电竞体育竞猜平台rdjournals.molbev.a040023
田村K。,Stecher G., Kumar S. (2021). MEGA11: Molecular evolutionary genetics analysis version 11.摩尔。杂志。另一个星球。38 (7),3022 - 3027。doi: 10.1093 / molbev / msab120
塔尔塔利亚C。,Azziz G。,Lorite M. J., Sanjuán J., Monza J. (2019). Phylogenetic relationships among introduced and autochthonous rhizobia nodulating三叶草种虫害在乌拉圭的土壤。达成。土壤生态。139年,40-46。doi: 10.1016 / j.apsoil.2019.03.014
托雷斯Tejerizo G。,Rogel m·A。,Ormeño-Orrillo E., Althabegoiti M. J., Nilsson J. F., Niehaus K., et al. (2016). Rhizobium favelukesii sp. nov., isolated from the root nodules of alfalfa (紫花苜蓿l。)。Int。j . @系统。Evolut。Microbiol。66 (11),4451 - 4457。doi: 10.1099 / ijsem.0.001373
托雷斯Tejerizo g . T。,德尔爸爸m F。,德拉吉W。,Lozano M。,Giusti M. de los Á., Martini C., et al. (2011). First genomic analysis of the broad-host-range根瘤菌sp。LPU83压力,较低的遗传多样性的一员Oregon-like根瘤菌sp.组。生物科技j . @》。155 (1),3 - 10。doi: 10.1016 / j.jbiotec.2011.01.011
托雷斯Tejerizo g . T。Wibberg D。,温克勒A。,Ormeño-Orrillo E., Martínez-Romero E., Niehaus K., et al. (2017). Genome sequence of the symbiotic type strain根瘤菌tibeticumCCBAU85039T。基因组的公告5 (4),e01513-e01516。doi: 10.1128 / genomeA.01513-16
韦格纳C。,Schröder S., Kapp D., Pühler A., Segundo López E., Martínez-Abarca F., et al. (2001). A population of twelve acid tolerant alfalfa nodulating根瘤菌菌株分离出不同的站点在阿根廷表现出相同的分子特征根瘤菌Or191 sp.压力。共生141 - 162年。
威尔逊K。,Sessitsch A., Corbo J., Giller K., Akkermans A., Jefferson R. (1995). B-glucuronidase (GUS) transposons for ecological and genetic studies of rhizobia and other gram-negative bacteria.微生物学141 (7),1691 - 1705。doi: 10.1099 / 13500872-141-7-1691
于G。,Smith D. K., Zhu H., Guan Y., Lam T. T. Y. (2017). Ggtree: an r package for visualization and annotation of phylogenetic trees with their covariates and other associated data.生态方法。另一个星球。8(1),几个。2041 - 210 - x.12628 doi: 10.1111 /
关键词:根瘤菌、Oregon-like应变、竞争力、苜蓿、M16肽酶
引用:Berais-Rubio,莫雷尔Revetria马,Gimenez M, Signorelli年代和蒙扎J(2023)竞争力和共生效率的紫花苜蓿根瘤菌favelukesiiORY1菌株的同源基因肽酶HrrP被确定和SapA负面影响的共生关系。前面。阿格龙。4:1092169。doi: 10.3389 / fagro.2022.1092169
收到:2022年11月07;接受:2022年12月02;
发表:2023年1月04。
编辑:
Basharat阿里,哈法里德·大学工程和信息技术(KFUEIT),巴基斯坦版权©2023 Berais-Rubio,莫雷尔Revetria,,他和蒙扎。这是一个开放分布式根据文章知识共享归属许可(CC)。使用、分发或复制在其他论坛是允许的,提供了原始作者(年代)和著作权人(s)认为,最初发表在这个期刊引用,按照公认的学术实践。没有使用、分发或复制是不符合这些条件的允许。
*通信:豪尔赫·蒙扎,jmonza@fagro.edu.uy
__ORCID:Andres Berais-Rubio,orcid.org/0000 - 0002 - 0259 - 9112
玛丽亚·a·莫雷尔Revetria,orcid.org/0000 - 0002 - 9064 - 5675
Matias,orcid.org/0000 - 0002 - 6267 - 9106
圣地亚哥Signorelli,orcid.org/0000 - 0002 - 1854 - 3164
豪尔赫·蒙扎,orcid.org/0000 - 0002 - 4309 - 7397