使用环肽抗原生成LRP8特定抗体
 玛丽亚a Argiriadi
1*__,
玛丽亚a Argiriadi
1*__, Kangwen邓1
__,大卫·伊根2,Lei高1,
Kangwen邓1
__,大卫·伊根2,Lei高1, 法Gizatullin
1,
法Gizatullin
1, 约翰·哈伦
2,丹尼斯Karaoglu Hanzatian1,魏邱2,
约翰·哈伦
2,丹尼斯Karaoglu Hanzatian1,魏邱2, 露丝维兰纽瓦
1和安德鲁Goodearl1
露丝维兰纽瓦
1和安德鲁Goodearl1
- 1AbbVie生物学研究中心,伍斯特,妈,美国
- 2AbbVie Inc .)、美国芝加哥,,
LRP8是LDLR-like蛋白家族的一员。这个运输受体,可用于抗体的设计具体的调查增加暴露疗法对血脑屏障(BBB)。在这项研究中,一个LRP8肽免疫策略是产生抗体来实现一个特定的抗原决定基LRP8 CR1领域的交叉反应,可以使传递函数和老鼠,猕猴和人类。此外,环化肽免疫原是为了保护结构β-hairpin元素之前解决了晶体结构中观察到一个相关的CR域。由于这种抗原设计小,LRP8特定的抗体,11个h1,选择和配体结合化验和晶体结构特征的决心。11个h1工厂的高分辨率结构复杂的环化CR1肽显示关键交互驱动抗原决定基承认使用定点诱变的方法被证实。关键的观察是,确定结构CR1抗原决定基11 h1没有与reelin承认CR1允许同时绑定。这是一个预测的在网上三元模型,并证实了reelin绑定数据。这些同时绑定事件(h1 / CR1 / reelin 11日)可以启用CR1 LRP8域,11个h1和reelin用作“BBB运输车”三元复杂设计的治疗性蛋白质。更重要的是,11个h1显示增强的大脑渗透全身静脉给药后小鼠的研究中,证实其势函数作为治疗性蛋白质BBB运输车。
1介绍
有许多抗体药物目前临床评估等神经系统疾病定位错误折叠蛋白清除阿尔茨海默病和帕金森病以及其他神经迹象,包括中风和更广泛的taopathies和痴呆类(Alpaugh Cicchetti, 2019)。共同挑战实现完整的功效在这些疗法的存在血脑屏障(BBB),限制暴露系统给蛋白质疗法在大脑的实质空间等靶细胞神经元和神经胶质驻留和其药理机制在玩(Terstappen et al ., 2021)。BBB的形成主要是由微血管内皮细胞周围的大脑形成的腔相互重叠,由健壮的紧密连接,连接在一起的周和星形endfeet。这种多细胞结构用来限制paracellular血管腔的水流和溶质包括金属离子、脂质、激素、血清蛋白质和抗体。许多薄壁组织的细胞运送营养至关重要通过特定的细胞在内皮细胞受体,可以绑定到的同源配体鲁米那(顶端)表面和内化和胞内运输后,释放实质(基底)表面上的配体通过transcytosis的过程。治疗生物工程药物的概念结合运输受体在大脑内皮细胞影响高架实质暴露(受体介导transcytosis (RMT)第一次证明了在活的有机体内使用特定的转铁蛋白受体抗体(Pardridge et al ., 1991)。从那时起,优化交通属性特征(转铁蛋白受体Yu et al ., 2011;Karaoglu Hanzatian et al ., 2018;Chang et al ., 2021)和增强在活的有机体内大脑吸收目标bi-specific生物制剂已经演示了额外的运输包括胰岛素受体,受体(Pardridge et al ., 1995)、CD98 (法灵顿et al ., 2014;Zuchero et al ., 2016)、IGF1R (Stanimirovic et al ., 2015)、FC5 (Abulrob et al ., 2005)和叶酸受体(吴和Pardridge 1999;Grapp et al ., 2013)。这些航天飞机目标与成功应用于临床主要在酶替代疗法空间(Terstappen et al ., 2021)。许多这些受体广泛表达在许多类型的细胞和组织,包括外围内皮细胞,这可能会限制其使用BBB运输受体。LRP8就是一个例外,也称为载脂蛋白e受体2,除了在血小板,选择性地表达在外围只有大脑内皮细胞。基于这种独特的表达谱,我们决定生成一组抗体LRP8更好地描述其潜力作为BBB运输车治疗蛋白质。
LRP8 LDLR-like蛋白家族的成员,是一个通过跨膜蛋白的氨基端细胞外域(ECD)由多个模块包括七LDLR类(LA)重复域,三个EGF重复使用嵌入式β-propeller域和一个O-linked糖基化域(但是,2009)。LRP8受可变剪接共有九个拼接变异识别在mRNA水平。除了作为受体对载脂蛋白E,进一步LRP8 ECD-binding配体已确定包括Reelin, (D 'Arcangelo et al ., 1999)、硒蛋白P (Kurokawa et al ., 2014),Clusterin (ApoJ)、血小板反应蛋白(布莱克et al ., 2008)和F-spondin (锄头et al ., 2005)。绑定在LRP8抗原表位建立了这些配体,包括LA1 (Reelin)和β-propeller域(硒蛋白P)。基因敲除小鼠的LRP8非致命,导致lissencephaly,条件的变化在大脑和小脑皮质折叠结构的发展导致受损的运动(黑客et al ., 2007)。这种表型非常类似于reelin击倒,暗示作用为主要reelin LRP8受体在开发过程中。成人LRP8功能尚不明朗,虽然它可能包括一个可能的ApoE代谢作用与其他家庭成员部队。
LRP8等细胞表面蛋白免疫原选项包括cDNA质粒,高表达细胞株重组,重组细胞外域和肽。作为并行免疫策略的一部分,我们评估肽免疫原的潜力。替代抗原肽免疫原形式,它可以是有利的蛋白质,因为他们可以可靠地准备了大量高纯度和稳定性。此外,序列可以选择特定的抗原决定基,例如通过选择一个同源序列区域生成与交叉反应的抗体靶蛋白在不同物种。为了提高交叉反应的几率本机折叠蛋白质的肽序列,蛋白质三维结构可以使用以模仿自然在特定地区三维目标蛋白质的构象。结构和热力学研究预测免疫的成功基于线性肽的稳定性(卡马乔et al ., 2008)。例如,某些二级结构元素可以有助于提高肽抗原包括β-turns和/或螺旋(李et al ., 2016)。虽然长肽可以采用这样的结构元素,环肽被用来以一种可预见的方式保存这些结构特点在短肽免疫接种和抗原抗体识别(三角et al ., 2003;Jakab et al ., 2009)。在这项研究中我们描述了使用环肽的免疫策略,导致鼠标/猕猴猴/人类可交叉反应的LRP8特定的抗体,11 h1。我们也使用配体绑定描述绑定抗原决定基的特征分析,晶体结构测定和建模分析与野生型和突变体肽。
2材料和方法
2.1创建的CR抗原肽
LRP8是一个大型的多畴的蛋白质。为了选择一个肽段作为一个健壮的抗原,我们最初指的是之前解决高分辨率结构(PDB 1.8代码1 j8e) CR7从人类的低密度脂蛋白受体相关蛋白(单体)(Simonovic et al ., 2001)。回顾,我们验证这种方法通过检查CR1域与小鼠reelin复杂(PDB代码:3 a7q) (Yasui et al ., 2010)。在LRP8密切检查结构和序列后,肽段选择模仿β-turn主题2β链组成的一个循环。为了稳定这个主题,肽环化被认为是。一个“孤”序列中的半胱氨酸突变Ser防止聚合。这种策略被用来改善蛋白质聚合与最近报道的例子(Saetang et al ., 2022)。由此产生的肽合成了新英格兰肽(加德纳,MA)(赛克(1.130)H2N-CEKDQFQSRNERCIPSVWR (KAoa)酰胺。类似的肽CR2还创建了基于结构对齐[赛克(1.130)]H2N-CADSDFTSDNGHCIHERWK (KAoa)酰胺。
2.2创建LRP8 11 h1.5b2抗体
KLH-conjugated肽内和由音乐厅被新英格兰合成肽(加德纳,MA)。等量的(50µg)内,由音乐厅是混合免疫和注射小鼠皮下注射每3周4次老鼠脾脏是收获。淋巴细胞分离和融合NS0细胞有根深蒂固的协议。杂种细胞上清液(SN)用于细胞流式细胞仪hLRP-8-HEK293稳定的细胞和父母HEK293细胞。上层清液(SN)绑定到hLRP-8-HEK293稳定的细胞,而不是父母HEK293细胞被选中。
2.3 LRP8稳定细胞系的一代
HEK293H细胞培养T25培养瓶和孵化37°C, 5%的公司2每4 - 5天,细胞通道。当天在转染前细胞稀释2×10 * 5细胞6板在99%细胞生存能力。智人(人类)低密度脂蛋白受体相关蛋白8 (LRP8) (isoform3)(加入# NP_059992)序列从基因库被确认。亩骶(鼠标)低密度脂蛋白受体相关蛋白8 (LRP8) (isoform2)(加入# NP_001074395)序列从基因库被确认。猕猴LRP8被确认在房子新创克隆。LRP8基因克隆到pCMV向量。2.5μg质粒DNA的混合和10μL Lipofectamine 2000(英杰公司)500年μL Opti-MEM孵化在室温20分钟,然后加入细胞。细胞在37°C 5%孵化有限公司24 h。4 h后,细胞培养在培养基37°C公司5%2过夜。在转染后的第二天,2毫升的选择媒体5毫克/毫升g - 418(最终浓度)补充道。增长媒体改变了转染细胞每隔4 - 5天。克隆稳定细胞系被连续稀释亲代细胞的生成和随后的孤立的单个细胞的扩张殖民地。描述细胞系的流式细胞仪,HEK293转染细胞被分离细胞使用生长介质,洗,re-suspended在寒冷的PBS (pH7.2) / 2%的边后卫(流式细胞仪缓冲区)1×10 e6细胞/毫升,孵化1 h在4°C的主要抗体和分析由Accuri C6。
2.4 ELISA
肽或重组蛋白涂在MSD 96 - (MSD猫# L15XB-3 / L11XB-3)板块和孵化一夜之间在4°C。盘子洗和阻止使用15%的边后卫(Hyclone,热科学猫# SH300700.03)在室温下与温和搅拌30分钟,盘子洗DPBS 3倍和抗体被添加。1 h(孵化后在室温下,盘子洗了DPBS又山羊反(MSD猫# R32AJ-1)或山羊anti-mouse磺基标记(MSD猫# R32AC-1)补充道。板在室温下孵化了1 h, DPBS清洗和沉浸在MSD读缓冲区(MSD猫# R92TC-2)在阅读之前MSD部门成像仪6000。使用Xlfit4软件包EC50值了。
2.5测量抗体亲和力(细胞MSD)
HEK293细胞overexpressing人类,猴子,或鼠标LRP8被添加到默沙东- 96 -孔板(MSD猫# L15XB-3 / L11XB-3)和孵化37°C 1 h。细胞被封锁使用15%的边后卫(Hyclone,热科学猫# SH300700.03)在室温下与温和搅拌30分钟,盘子洗DPBS 3倍和抗体被添加。1 h孵化后在室温下,盘子洗了DPBS又山羊反(MSD猫# R32AJ-1)或山羊anti-mouse磺基标记(MSD猫# R32AC-1)补充道。板在室温下孵化了1 h, DPBS清洗和沉浸在MSD读缓冲区(MSD猫# R92TD-2)在阅读之前MSD部门成像仪6000。使用Xlfit4软件包EC50值了。
2.6细胞流式细胞仪
LRP8稳定细胞和父母HEK293细胞收集和孵化流式细胞仪缓冲区(1 xpbs + 2% fcs),和一个整除控制井被拔掉。亲代细胞与CFSE标记(5 (6)-Carboxyfluorescein N-hydroxysuccinimidyl酯)。50000细胞/的相同数量的CFSE标记亲代细胞和un-labeled LRP8稳定细胞混合,在1200转离心10分钟。添加流式细胞仪缓冲之后,细胞5×104细胞每口井被旋转2000转3分钟,然后孵化15分钟在4°C马伯从30μg /毫升和整个板的三倍。三洗后用流式细胞仪缓冲区,细胞被孵化15分钟在4°C 50μL二级热电电子APC抗体稀释1:50 0。无限二级抗体与流式细胞仪被三个洗缓冲区,然后细胞resuspended章50μL流式细胞仪和分析的缓冲区(BD FACSCanto)。
2.7突变分析
基于肽内和由音乐厅的排列顺序序列FxSxN似乎是常见的在这两种结合肽,因此可能重要的抗原决定基有约束力。注意,丝氨酸肽序列内,由音乐厅已经改变了从父母LRP-8蛋白质序列,删除一个未配对的半胱氨酸。修改形式的环肽CR1、包含所示的更改图8合成,检查绑定嵌合抗体(胡IgG1 / k) anti-LRP-8-11H1.5B2拉拉直接ELISA检测(图8 b)。化验,肽CR1.2 CR1.3绑定和亲和力的类似修改的内,而肽CR1.1 & CR1.4都包含F - >替换,没有绑定。这表明,苯丙氨酸sequence-FQSRN——所需的抗体绑定,因此残留在抗原决定基的关键。
2.8 11 h1.5b2工厂准备和净化
Fab片段LRP-8 11 h1.5b2准备了木瓜蛋白酶的乳沟父母抗体,抗-单体- 8.11 - h1.5b2(μ/胡锦涛IgG1 / k)拉拉嵌合抗体。木瓜蛋白酶与50 mM激活半胱氨酸在PBS, pH值7.4缓冲区。反-单体- 8.11 - h1.5b2嵌合抗体在PBS, pH值7.4缓冲和木瓜蛋白酶在1:10 0的重量比木瓜蛋白酶抗体和孵化1 h在37°C。用5毫米碘乙酰胺的反应就熄了。混合物的提纯是5毫升马伯SelectSure树脂(通用电气医疗集团)流过的Fab片段收集。流经集中使用以下Ultrafree-15 10 kDa分子量截止(MWCO)离心设备(微孔)。集中混合物纯化于200年2.6厘米×60厘米Sephacryl HiPrep列(通用电气医疗集团)在50 mM消息灵通的预平衡,50 mM氯化钠,pH值7.5缓冲区。
2.9结晶LRP8 11 h1.5b2复杂LRP8环肽
环肽与蛋白质溶解缓冲区(50毫米氯化钠,50毫米玫瑰,pH7.5)的最终浓度100毫米。添加肽是蛋白质(29.4毫克/毫升)的最后一个8:1的摩尔比(肽/蛋白)。堆叠薄板晶体最初观察2 - 3天之后。他们成长为全尺寸的条件下一周内25% PEG 4000, 0.2硫酸铵,0.1米/乙酸钠盐酸,pH值4.6。板晶体被分开,和flash冻成液态氮用20%丙二醇+水库cryo-protectant解决方案。衍射数据收集在100 K温度使用beamline XALOC ALBA同步加速器(BL13),西班牙。
2.10数据收集和嵌合的结构测定11 h1多元LRP8 CR1肽
x射线衍射数据收集11 h1 / CR1肽复杂晶体在阿尔巴同步beamline 1.72决议。晶体是维持在100 k的牛津Cryosystems Cryostream冷却器在数据收集。被处理的数据使用程序AUTOPROC从全球定相(Vonrhein et al ., 2011)。总结了x射线衍射数据和细化统计数据表1。以下列表索引的晶体形式:空间群P21= 41.3,67.1 b = 79.8 a, c =β= 95.5。
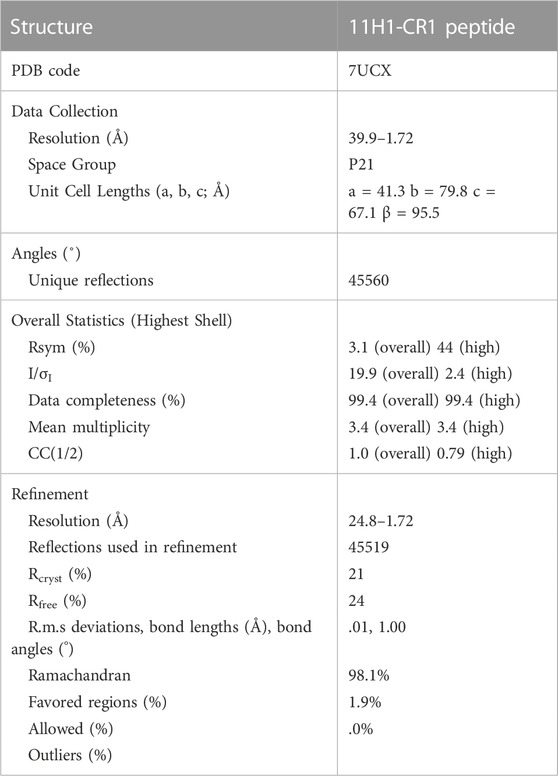
表1。晶体统计11 h1 / CR1肽复杂结构。
最大似然分子替代解决方案是决定使用程序移相器(McCoy et al ., 2007)使用工厂搜索模型之前报道(蛋白质数据库条目1 vpo, (Valjakka et al ., 2002)。坐标是基于分子生成的替代解决方案。产生的解决方案进行了初步改进使用REFMAC (Murshudov et al ., 2011)和程序巴斯特(布兰科et al ., 2004)。迭代的蛋白质模型建立工作使用傻瓜(Emsley卡谭,2004)检查2 Fo-Fc Fo-Fc电子密度地图。CR1肽是手动内置电子密度。过去的五肽的残留未见的电子密度可能由于这个区域的灵活性。此外,Cys1之间的交互和Cys13建模为二硫化是因为附近的蛋白质骨架,尽管该地区较弱的密度。细化得出的水分子使用巴斯特和分析程序套件凤凰(亚当斯et al ., 2010)。最后细化统计所示表1。
进行了分子动力学模拟的程序德斯蒙德(薛定谔释放2021 - 1:德斯蒙德分子动力学系统,d . e . Shaw的研究中,纽约,纽约,2021年。Maestro-Desmond互操作性工具,薛定谔,纽约,纽约,2021年。薛定谔/ DE Shaw)。20 ns仿真使用OPLS3e力场是由300 k。轨迹检查来确定智慧化。d为侧链和骨干运动范围。
2.11细胞reelin-anti-LRP8竞争分析
HEK293细胞overexpressing猕猴猴LRP-8被添加到默沙东- 96孔板(MSD猫# L15XB-3 / L11XB-3)和孵化37°C 1 h。细胞被封锁使用15%的边后卫(Hyclone,热科学猫# SH300700.03)在室温下与温和搅拌30分钟,盘子与DPBS洗3次。Anti-LRP-8抗体与人类Fc和reelin-His-FLAG被用于两个竞争组合:
竞争1(竞争对手:Anti-LRP8 Ab): 1:1的混合固定集中reelin-HIS-FLAG和竞争对手的效价含碘- 8.11 - h1或控制抗体被添加到板和孵化室温1 h。洗后,reelin绑定信号检测与anti-his Ab (Novex)。2的竞争(竞争对手:Reelin): 1:1的混合固定集中anti-LRP8.11H1 Ab和效价的竞争对手Reelin被添加到板和孵化室温1 h。洗后,添加了反SULFO-TAG Ab板和孵化1 h。洗了盘子DPBS和沉浸在MSD读取缓冲区T自由表面活性剂(MSD猫# R92TD-2)在阅读之前MSD部门成像仪6000。数据获取和分析使用GraphPad棱镜6软件包(GraphPad软件公司,拉霍亚,CA)。
2.12在活的有机体内研究
野生型女性C57Bl / 6 n小鼠(6 - 8周大)从泰康利购买生物科学,Inc .老鼠维护和处理按照下列动物保健机构和用户委员会(IACUC)批准的协议。每组四个动物注射50毫克/公斤的抗体通过静脉注射的路线。表示时间(24小时)后,动物安乐死的过量Ketamine-Xylazine (anas道奇堡)管理通过腹腔内注射。右心房是雕刻的,动物是transcardially灌注冷杜尔贝科的磷酸缓冲盐含有肝素(1000单位/ L) 2毫升/分钟的速度10分钟通过可编程的蠕动泵(ne - 1000)。血清和组织收集。
2.13测量老鼠组织和血清中抗体浓度
大脑解剖灌注从每个鼠标,垂直分成相等的两半;免疫组织学得救了一半,另一半是均质使用子弹搅拌机蓝色(NextAdvance BBX24B)和锆珠(NextAdvance, ZROB05 / ZROB10)在1% NP-40(热科学猫# 28324)PBS含有蛋白酶抑制剂(罗氏诊断完成迷你,EDTA-Free Ref # 11836170001)。均质大脑样本在4°C 1 h旋转旋转20分钟的14000 rpm。上层清液分离和抗体测量大脑使用ECL-MSD化验。
全血收集从心脏穿刺在终端的时间点。整个血液从心脏穿刺收集血清中分离器BD microtainer™管(双相障碍诊断,裁判# 365956),允许凝块30分钟,旋转在13000 rpm在室温下8分钟。上层清液分离和血清中抗体测定使用ECL-MSD化验。
在小鼠血清抗体浓度和组织样品测定ECL-MSD化验。默沙东- 96 -孔板(MSD猫# L15XB-3 / L11XB-3)被涂上一层F (ab) 2片段的驴反人类免疫球蛋白Fc fragment-specific多克隆抗体(杰克逊ImmunoResearch代码# 709-006-098)在一夜之间2μg /毫升4°C。盘子被封锁3% MSD阻断缓冲区(MSD猫# R93BA-04) 1 h 25°C。盘子洗了三次1 x Tween-Tris缓冲盐水,洗板机(ELx45 Bio-Tek仪器Inc .)。标准是由连续稀释1% MSD分析缓冲区或1% MSD分析缓冲区包含物质血清。组织样本稀释1:2或1:4的1%并分析缓冲区和血清样本连续从1:10稀释1% MSD分析缓冲和25μL(副本)被添加/。每个抗体作为内部标准量化各自的抗体浓度。盘子被孵化2 h 25°C和结合抗体检测与山羊反Sulfo-TAG (MSD猫# R32AJ-1)。盘子在MSD部门成像仪6000读。浓度测定的标准曲线有四个参数物流(4 pl)非线性回归程序从idb XLfit®微软®软件的插件。 The ECL-MSD assay lower limit of quantitation values ranged from .05–.46 ng/mL in serum and tissue samples. Molar concentration was calculated by the MSD quantification value (in ng/ml) that best fit its corresponding standard curve with a coefficient of variance ≤20% and within acceptable recovery of 80%–120% divided by the respective antibody molecular weight. For tissue samples, multiplication of tissue homogenate dilution factor was considered. Data was expressed as means ± standard deviation (SD) and statistics were assessed by unpaired, two-tailed t-tests. Total protein concentrations in brain extracts were measured using the BCA protein assay kit (Thermo Scientific, Cat#23225) and were consistently found to be within 15% of coefficient of variation (CV). Samples that did not meet these criteria were not used for analysis.
2.14免疫组织化学方法和分析
一半大脑从灌注antibody-treated老鼠浸固定在4%多聚甲醛固定后6 h。,组织通过一系列分级冲加工协议(徕卡TP1050组织处理器)的酒精二甲苯,然后嵌入在石蜡(徕卡EG1150H)。(5μm)大脑部分被剪掉了,切片机(Microm HM355S)。部分是de-paraffinized和水化水,放入三羟甲基氨基甲烷与Tween-20缓冲液(Teknova猫# T5155)。染色当时上执行一个Dako autostainer链接48系统。短暂的部分被封锁,3%过氧化氢和甲醇为30分钟,用10倍三羟甲基氨基甲烷与Tween-20缓冲液(Teknova猫# T5155)培养与蛋白酶8分钟我(Ventana Ref # 760 - 2018)。部分被封锁链霉亲和素与生物素阻断工具包(向量实验室猫# sp - 2002) 8分钟,其次是Dako蛋白质块30分钟。接下来,部分被孵化1 h与生物素化的驴在室温下反人类免疫球蛋白(h + L) F (ab)(杰克逊ImmunoResearch代码# 709-066-149)在15μg /毫升紧随其后的孵化与过氧化物酶共轭抗生物素蛋白在室温下30分钟(保留时间U ABC工具包(向量pk - 7100)]。部分被反应与diaminobenzidine (DAB)发色体(Dako Ref # K3468) 3分钟形成棕色沉淀,用水洗,与吉尔修改苏木精复染色(EMD Harleco Ref # 65065) 30年代和发蓝处理试剂浸幻灯片5 - 6次水库(richard allan科学裁判# 7301),脱水和安装显微镜观察。从四个不同的大脑区域部分(前脑、中脑后脑,小脑)从每组四个动物被染色。代表染色图像被奥林巴斯BX43或幻灯片和全景250幻灯片扫描仪扫描。所有设置(光过滤器和水平)为每个图像都是在整个实验过程中保持不变。
3的结果
3.1创建CR1抗原肽
LRP8是一个大型的多区域蛋白包含八个配体结合区域和一个含有半胱氨酸EGF-like域丰富的重复。为了选择一个LRP8肽段作为一个健壮的抗原,可用单体结构信息。1.8之前解决结构(PDB代码1 j8e) CR7从人类的低密度脂蛋白受体相关蛋白肽(含)成为一个起点工程(Simonovic et al ., 2001)。序列比对的LRP8 CR1、CR2的x射线结构1 j8e进行(图1一个),使肽段的设计将满足几个条件:1)不超过20肽残留2)肽序列是保存在多个直接同源如鼠标和cyno 3)肽抗体识别的足够的二级结构。

图1。环肽内和循环由音乐厅的设计。(一)序列比对的complement-like重复区域(CR1/2)域CR7 LRP8与晶体结构的单体(PDB代码1 j8e)。(B)晶体结构的CR7单体(PDB代码1 j8e)。骨干彩色和环绕蓝色是抗原肽的序列选择。(C)序列的两个CR肽合成免疫与特定C→S变异显示为红色。
仔细检查后1 j8e结构(图1 b肽段)和序列在LRP8,被选为CR1模仿β-turn主题β链之间的循环可以用于抗体识别。为了稳定这个主题,肽环化建议。此外,一个“孤”序列中的半胱氨酸改为Ser防止聚合生产过程中肽。基于对齐,CR2肽也是设计。结果合成肽所示图1 c。
3.2代anti-LRP8抗体使用肽内和由音乐厅作为免疫原
4 Balb / c和4 SJL小鼠免疫与皮下注射等量的KLH-conjugated肽内和由音乐厅(50µg)混合物。从小鼠血清测试通过流式细胞仪使用HEK293-LRP8-HA稳定细胞系和展出一些特异性(数据未显示)。2 SJL老鼠融合。杂种细胞上清液(SNs)被CR1筛查三轮,CR2肽ELISA(数据未显示)。选择的SNs, 11个h1, 20 e8和12 f6,肽ELISA和测试的3 - 81倍稀释在细胞流式细胞仪绑定化验使用LRP8表达HEK293细胞和父母HEK293细胞(图2)。SN 11 h1显示好绑定LRP8表达HEK293细胞(图2一个),而不是父母HEK293细胞(图2 b)。11个h1是最好的粘合剂和subcloned获得11 h1.5b2克隆。Anti-LRP8.11H1.5B2是鼠标IgG1 kappa轻链。11 h1.5b2的产量杂种细胞达到198 mg / L。纯度100%单体分子的尺寸排阻色谱(SEC)。

图2。小鼠免疫使用环肽内和由音乐厅。小鼠免疫与等量的KLH-conjugated肽内和由音乐厅。细胞流式细胞仪绑定1:3系列稀释的杂种细胞上清液(SN)(一)hLRP-8-HEK293稳定细胞和(B)父母HEK293细胞了。Anti-HA是积极控制抗体。颜色代表了连续稀释系数。
可变域得到11 h1.5b2 cDNA克隆序列。嵌合anti-LRP8.11H1.5B2用鼠标可变域和人类恒域构造和生产是暂时性的转染HEK293细胞。纯化anti-LRP8.11H1.5B2 m / hIgG绑定到肽1.5和2.1 nM内和由音乐厅亲和力,分别为(图3)。序列之间的同源性鼠标/ cyno /人类LRP8以下:人类与老鼠89%,人类与cyno 86%。域内和由音乐厅之间的序列同源性为50%。在细胞MSD绑定化验使用LRP8-overexpressing HEK293细胞,11 (h1绑定到老鼠。cyno(27海里)。46nM) and human (.32 nM) LRP8 with high and comparable affinity (图3 b)。的最大约束力的信号差异是由于LRP8表达水平这些细胞系。控制hIgG没有绑定。另外,11个h1证明没有绑定的C2和C3域LRP1恰巧,这是另一个单体受体家族的成员,其中包含相关CR域(图3 c)。

图3。Anti-LRP8.11H1绑定两个CR1、CR2肽和LRP8表达细胞。(一)纯化anti-LRP8.11H1抗体结合两种肽内和由音乐厅。(B)Anti-LRP8.11H1抗体或控制免疫球蛋白结合HEK293细胞过表达人类LRP8 cyno LRP8或鼠标在细胞LRP8 MSD化验。(C)Anti-LRP8.11H1结合具体重组人类LRP8 (rhLRP8)和不绑定到rhLRP1_C2和rhLRP1_C3域。同形像统计图控制抗体显示最少的绑定。
3.3晶体结构的11个h1外事局多元CR1肽
11 h1工厂复杂的晶体结构CR1肽是解决分辨率(1.7图4)。结构解决了用分子替换先前报道的晶体结构(PDB: 1 vpo) (Valjakka et al ., 2002)。大部分工厂的电子密度和肽是明确的。只有5 c端肽残留并不适合密度由于蛋白质的灵活性。肽被发现之间的绑定到11个h1工厂工厂的轻、重链与多个联系人界面。此外,CR1肽假设循环构象,由一个稳定的二硫桥近距离预测的建模Cys1和Cys13 (图5一个)。这二硫桥也观察到在前面描述的x射线结构,1 j8e (Simonovic et al ., 2001)。当覆盖在工厂看到的肽结构与最近报道reelin CR1结构复杂(PDB代码:3 a7q,图5 b)(Yasui et al ., 2010),二硫循环结构相似的循环结构有关。然而,工程肽假定两个螺旋与β链3 a7q结构观察。启用了意想不到的螺旋转几个关键残留项目进入工厂轻、重链界面。几个关键的相互作用参与了11个h1 Fab-LRP8 CR1抗原决定基。肽侧链从Phe6 Gln5插入口袋里,参与多个交互如分子动力学2 d输出(图6)。在分析x射线结构,CR1肽Gln5与周围的水分子进而氢连着的侧链(L) Tyr41和Ser94 (L)。Gln也是近端(H)的sidechains Thr50 (H) Ser35但不直接参与氢键相互作用。肽的骨干NH Gln5与(H) Asp99直接交互。CR1肽Phe6了edge-to-faceπ−π相互作用(H) Trp47除了芳香sidechain的包装(H) Tyr59 (图7)。重要的是要注意,CR2还包含一个板式换热器在这个位置,也可能包含这些残留物。额外的关键CR1肽之间的氢键相互作用观察Glu2 (H) Ser52和CR1肽Asp4 (H) Tyr59 (图7 b)。

图4。晶体结构的11个h1外事局多元CR1肽。轻链是彩色的绿色(左蛋白质链),重链是彩色tan(右蛋白质链)。轻、重链之间的肽位于CDR的显示和由表面。

图5。放大的视图11 h1工厂多元CR1肽。覆盖的x射线肽构象和CR1 LRP8。(一)。轻链是彩色的绿色(左蛋白质链),重链是彩色tan(右蛋白质链)。肽假设循环构象(中心)所示通过二硫键从终端的肽。(B)环肽(红色圆圈)假定的构象,类似于自然循环LRP8 CR1脚手架(从PDB代码3 a7q紫色),然而肽假定一个独特的螺旋构象在x射线结构。

图6。MD CR1绑定到抗体的分析。20 ns进行了分子动力学模拟的11个h1 / CR1肽结构程序德斯蒙德在这个模型使用OPLS3e力场300 K和pH值7.0。交互观察作为课程的时间一致的关键Phe6和Gln5肽残留在工厂间隙。

图7。特定的相互作用CR1肽的轻、重链界面11 h1。(一)CR1肽Gln5(红色)与水分子相互作用进而氢连着的侧链(L) Tyr41和Ser94 (L)。CR1肽残渣Phe6(红色)演示了一个π−π芳香互动(H) Trp47除了显示接近Tyr59 (H)。(B)额外键之间的交互观察CR1肽Glu2 (H) Ser52和CR1肽Asp4 Tyr59 (H)。
确认的重要性肽Phe6残渣,正交定点诱变方法进行。四个突变体变异CR1肽合成的几个潜在的11个h1互动氨基酸苯丙氨酸(F)丝氨酸(S)和天冬酰胺丙氨酸(A) (N) (图8)。11个h1结合肽是迷失在苯丙氨酸(F)突变体CR1.1 CR1.4和不受S N突变CR1.2和CR1.3 (图8 b)。

图8。突变分析表明,苯丙氨酸(F)是绑定的关键氨基酸。(一)肽CR1.1、CR1.2 CR1.3和CR1.4 CR1的丙氨酸突变体。(B)CR1.2和CR1.3结合亲和力的类似修改的内,而肽CR1.1 & CR1.4都包含F→一个替换,没有绑定到anti-LRP8.11H1。这两条曲线直接重叠。
当比较在序列内,由音乐厅,它是假定二硫化的关键桥梁被保存为指出这两个序列的保守的半胱氨酸。图1一个显示了两个域的序列比对。近端,该抗原法关键Gln在CR2 CR1是Asp,这提出了一个排斥向附近(H) Asp99 11 h1。这可能解释为什么11 h1不作为一个活页夹的紧CR2 CR1。此外,在CR1、先于Gln的Asp CR1 (H) Tyr59关键交互。这个职位是CR2的爵士,这可能是太短让(H) CDR2这种交互。
3.4分子动力学与reelin 11 h1-cr1复杂的和整体三元复杂
为了证实肽的稳定性绑定在抗体绑定崩裂,一个20 ns分子动力学模拟是在项目进行的德斯蒙德(薛定谔释放2021 - 1:德斯蒙德分子动力学系统,d . e . Shaw的研究中,纽约,纽约,2021年。Maestro-Desmond互操作性工具,薛定谔,纽约,纽约,2021)。水分子的溶剂化模型,进行了仿真在300 k pH值7.0 OPLS3e使用力场。在仿真的过程中大多数CR1肽的相互作用和11个h1是通过集群维护的轨迹。有趣的是,MD输出显示的潜在重要性芳香族侧链间的相互作用肽Phe6和Tyr59 (H)。二硫键的形成环肽结构,也是维护结构。此外,Cα骨干显示稳定RMSF范围。6 - 2.0之间。
Reelin 5、6片段包括Reelin单体绑定域(Yasui et al ., 2010)。当比较表面的交互LRP8 CR1 reelin CR1肽的相互作用表面和11 h1.5b2工厂,关键11 h1.5b2肽识别残留Asp-Gln-Phe并非用于reelin识别,这表明reelin-CR1-1H11.5B2三元可以实现。图9这三元复杂的显示生成的模型,在大师创建使用前面决定3 a7q结构和新的11 h1复杂结构作为模板。使用蛋白质制备三元复杂进一步最小化工作流的大师。模型中观察到,工厂的轻、重链抗体没有sterically对阵reelin支架允许同时使用CR1域蛋白质-蛋白质之间的关系作为一个双重“运输车”约束力的合作伙伴。这个模型同意reelin绑定数据图9 b, C。具体来说,这个数据显示reelin R5, 6-His-FLAG绑定到LRP8表达HEK293细胞。此外,anti-LRP8.11H1没有竞争reelin绑定在细胞LRP8竞争分析与固定365 nM reelin和滴定11 h1 10μg /毫升,或固定2 nM的11个h1和滴定reelin 365海里。

图9。11个h1和Reelin CR1绑定到相反的一面。(一)的三元模型Reelin多元CR1和11个h1。(B, C)基于单元的竞争分析与LRP8表达HEK293细胞涂井和孵化与不同比例的混合物reelin和11个h1。11个h1不会干扰reelin绑定,支持模型。
3.5 11 h1抗体显示更好的渗透到老鼠的大脑
为了证实anti-LRP8.11H1抗体的大脑渗透功能,50毫克/公斤anti-LRP8.11H1抗体或同形像控制抗体注入C57BL / 6小鼠静脉注射。11个h1血清中浓度是2379±473海里,这是与同形像控制抗体(1741±924海里)。11个h1的老鼠的大脑吸收水平2倍相比,同形像控制抗体。小鼠大脑组织的免疫组织化学染色显示良好的血管染色的大脑区域。增强的实质染色相比,所有的大脑区域控制观察免疫球蛋白。神经元染色在脑桥髓质和脊髓也观察到(图10)。

图10。增强小鼠大脑吸收anti-LRP8.11H1控制免疫球蛋白抗体相比。(一)免疫组织学染色老鼠大脑部分注入静脉后24 h抗体。皮质部分与脉管系统(黑色箭头)和实质染色与神经元和脊髓节染色(红色箭头)。(B)抗体浓度在24 h后单一静脉注射50毫克/公斤显示显著增强大脑吸收相比,同形像控制抗体。数据被表示为%的大脑/血清,n= 4老鼠每组。
4讨论
在这项研究中,抗体生成,公认的循环CR1肽和全长LRP8 CR1域。这表明1)这一领域的蛋白质,为设计选择,访问了抗体绑定和2)肽认为循环构象,模仿一个抗原循环地区土生土长的蛋白质。抗原肽的设计使用了一个基于结构的方法加上仔细观察CR域序列满足必要的条件来实现一个健壮的LRP8抗原决定基。直接同源大而稳定的结构主题是研究早期CR结构时的首要考虑。尽管CR域的体积小,有几个区域检查,这是一个二硫稳定地区的这个领域优先的环肽的设计支持LRP8杂种细胞运动。因此,选择性LRP8试剂,11个h1,在绑定化验跨物种识别和确认。结构研究证实了最初的假设,一个孤立的结构图案可以作为抗原识别的一种手段。此外,定点诱变证实抗原肽热点残基的重要性。分子建模和动力学实验揭示,不仅工厂/ CR1识别的稳定性,而且固化要求LRP8可以同时绑定reelin和抗体。因此,结合实验和在网上肽设计被证明是非常成功的大型蛋白质如LRP8锻炼。通过这个工作流,我们能够专注于一个小但健壮的领域追求的蛋白质直接同源大和双reelin BBB运输所需的绑定。一般来说,应该实现结构性方法通常在设计稳定、独特,目标蛋白质的二级结构元素,是一种强大的方法来创建抗原进行杂种细胞活动。还有其他技术,可以考虑这种类型的设计方法。回顾历史,当想到我们的结果有可能考虑策略的改进。
的肽设计、reelin复杂结构没有可供检查。一个考虑因素是证实了我们最初的建模/设计假设和选择的整个CR1肽通过定点突变技术领域对reelin绑定数据。变异位置,不会影响reelin绑定,将优先肽结合。第二个考虑的是确认的二级结构正交技术,如圆二色性(CD)。这种技术被识别螺线卷曲转换肽(问题et al ., 2014)。我们的结构表明,CR1肽假定两个螺旋转11 h1裂,这是不同的构象CR1的释放形式。监测二级结构通过CD不同肽可能是一种手段的优化。额外的考虑是解决整个CR1的co-crystal结构域有11个h1工厂,这将1)确认螺旋抗原二级结构的全域和2)提供额外的结构信息来优化肽的设计。
除了二硫桥接肽稳定,有额外的战术,这可能是用于驱动二级结构:交联使用内酰胺桥接或thiol-reactive alkyne-based cross-linkers (欧洲蕨et al ., 1994;Zhang et al ., 2007)。这些方法已经使用了一些蛋白质的目标。观察11 h1的螺旋形识别后,钉肽的方法可能是一个适当的后续策略。例如,烃装订与gp41演示特定的肽,以制定疫苗接种hiv - 1免疫原。这种装订方法辅助保留生物活性一个肽的螺旋构象(鸟et al ., 2014)。搭配CD分析,所有这些不同的连接方法可能导致进一步优化的生物活性肽。
最后一个考虑是比较LRP8 CR1序列与其他家庭成员需要考虑大和备用所需的绑定配置文件。例如,当检查序列和结构模型比较,LRP1恰巧有一个相同的苯丙氨酸,是至关重要的接口LRP8 CR1-11H1结构(图11 a, B)。然而正如前面所提到的,LRP1恰巧肽包含不同的外围残留物,改变静电表面轮廓的肽当假设循环构象(图11 c)。如果期望的结果是创建一个交叉反应抗体LRP1恰巧和其他亚型的家庭,静电分析除了设计多肽与序列的比较将是必要的。通过初步序列检验、序列之间的身份LRP1恰巧CR1和LRP8 CR1是c端观察序列的一部分。肽设计这部分的序列可能导致更高的大。

图11。(两者)序列、结构和静电比较CR1肽LRP1恰巧(左)和LRP8(右)。
尽管它曾推断序列同源性,LRP8可能交通属性,在这个报告我们实验证实,LRP8可以增强大脑的BBB目标绑定蛋白质的吸收利用RMT通路。的大脑吸收增加anti-LRP8.11H1tool抗体是温和的(2 - 3倍),但重要的是要注意,绑定关联可能不是最优的,可能是由于其相对较高的亲和力LRP8。其他BBB RMT目标,它已经表明,大脑吸收的高亲和力抗体可以改善通过减少其亲和力BBB目标(Yu et al ., 2011;Karaoglu Hanzatian et al ., 2018)。演示功能BBB航天飞机属性利用大脑内皮细胞的具体表达式LRP8 (Zhang et al ., 2014;Munji et al ., 2019;Zhang et al ., 2020)等待进一步的研究关联优化LRP8 bi-specific分子。
最后,我们使用一个基于结构的方法来设计一个目标LRP8肽抗原为BBB运输。产生的抗体是一种选择性试剂对其他家庭成员部队由于抗原决定基的关键交互识别。此外,LRP8 CR1域可以同时绑定到这个抗体及其约束力的合作伙伴,reelin,使reelin之间的“交通”三元复杂,LRP8和11个h1。总的来说,这突显出探索基于结构的设计可以提高抗原肽的创造和增加成功识别可靠的生物试剂为目标蛋白质的兴趣。
数据可用性声明
PDB数据提出了研究可以发现在网上存储库https://www.rcsb.org加入以下代码:7 ucx。
道德声明
动物研究是审查和批准机构动物保健和用户委员会(IACUC)。
作者的贡献
马、KD和AG)准备手稿。妈,KD、JH WQ, AG)、德、LG、FG,房车,DKH所有参与评审的手稿。妈,KD、JH WQ,德,LG,房车,FG提供实验数据支持这个手稿。
确认
作者感谢夜巴洛(前AbbVie雇员)有价值的科学讨论关于LRP8 11 h1项目。
的利益冲突
DKH MAA, KD、LG、FG,房车和AG是受雇于AbbVie生物学研究中心。德,JH, WQ受雇于AbbVie Inc .)
设计、研究行为,AbbVie提供的金融支持这项研究。AbbVie参与解释数据,审查,批准发布。
出版商的注意
本文表达的所有索赔仅代表作者,不一定代表的附属组织,或出版商、编辑和审稿人。任何产品,可以评估在这篇文章中,或声称,可能是由其制造商,不保证或认可的出版商。
引用
Abulrob,。,Sprong, H., Van Bergen en Henegouwen, P., and Stanimirovic, D. (2005). The blood-brain barrier transmigrating single domain antibody: Mechanisms of transport and antigenic epitopes in human brain endothelial cells.j . Neurochem。95 (4),1201 - 1214。doi: 10.1111 / j.1471-4159.2005.03463.x
亚当斯,p D。,Afonine, P. V., Bunkoczi, G., Chen, V. B., Davis, I. W., Echols, N., et al. (2010). Phenix: A comprehensive python-based system for macromolecular structure solution.Acta Crystallogr。d杂志。Crystallogr。66 (2),213 - 221。doi: 10.1107 / S0907444909052925
Alpaugh, M。,和Cicchetti, F. (2019). A brief history of antibody-based therapy.一般人。说。130年,104504年。doi: 10.1016 / j.nbd.2019.104504
问题,K。,Liyanage, M. R., Volkin, D. B., and Middaugh, C. R. (2014). Circular dichroism of peptides.摩尔。生物方法。1088年,247 - 253。doi: 10.1007 / 978 - 1 - 62703 - 673 - 3 - _17
鸟,g . H。,Irimia, A., Ofek, G., Kwong, P. D., Wilson, I. A., and Walensky, L. D. (2014). Stapled HIV-1 peptides recapitulate antigenic structures and engage broadly neutralizing antibodies.Nat。结构。摩尔。杂志。21 (12),1058 - 1067。doi: 10.1038 / nsmb.2922
布莱克,s M。,Strasser, V., Andrade, N., Duit, S., Hofbauer, R., Schneider, W. J., et al. (2008). Thrombospondin-1 binds to ApoER2 and VLDL receptor and functions in postnatal neuronal migration.EMBO J。27日(22日),3069 - 3080。doi: 10.1038 / emboj.2008.223
布兰科,E。,Roversi, P., Vonrhein, C., Flensburg, C., Lea, S. M., and Bricogne, G. (2004). Refinement of severely incomplete structures with maximum likelihood in BUSTER-TNT.Acta Crystallogr。d杂志。Crystallogr。60 (1),2210 - 2221。doi: 10.1107 / S0907444904016427
欧洲蕨,C。,Gulyas, J., Taylor, J. W., and Baum, J. (1994). Synthesis and nuclear magnetic resonance structure determination of an .alpha.-Helical, bicyclic, lactam-bridged hexapeptide.j。化学。Soc。116 (14),6431 - 6432。doi: 10.1021 / ja00093a052
卡马乔,c·J。,Katsumata, Y., and Ascherman, D. P. (2008). Structural and thermodynamic approach to peptide immunogenicity.公共科学图书馆第一版。医学杂志。4 (11),e1000231。doi: 10.1371 / journal.pcbi.1000231
常,h . Y。吴,S。李,Y。,Zhang, W., Burrell, M., Webster, C. I., et al. (2021). Brain pharmacokinetics of anti-transferrin receptor antibody affinity variants in rats determined using microdialysis.马伯13 (1),1874121。doi: 10.1080 / 19420862.2021.1874121
D 'Arcangelo, G。Homayouni, R。Keshvara, L。、大米、d S。谢尔登,M。,和Curran, T. (1999). Reelin is a ligand for lipoprotein receptors.神经元24 (2),471 - 479。doi: 10.1016 / s0896 - 6273 (00) 80860 - 0
Emsley, P。,和Cowtan, K. (2004). Coot: Model-building tools for molecular graphics.Acta Crystallogr。d杂志。Crystallogr。60 (1),2126 - 2132。doi: 10.1107 / S0907444904019158
法灵顿·g·K。Caram-Salas, N。哈卡尼,a。黑发,E。、·J。,Pepinsky B。,et al. (2014). A novel platform for engineering blood-brain barrier-crossing bispecific biologics.美国实验生物学学会联合会J。28 (11),4764 - 4778。doi: 10.1096 / fj.14 - 253369
Grapp, M。,Wrede, A., Schweizer, M., Huwel, S., Galla, H. J., Snaidero, N., et al. (2013). Choroid plexus transcytosis and exosome shuttling deliver folate into brain parenchyma.Commun Nat。4、2123。doi: 10.1038 / ncomms3123
黑客,我。,Hellwig, S., Junghans, D., Brunne, B., Bock, H. H., Zhao, S., et al. (2007). Divergent roles of ApoER2 and Vldlr in the migration of cortical neurons.发展134 (21)3883 - 3891。doi: 10.1242 / dev.005447
锄头,h·S。,Wessner, D., Beffert, U., Becker, A. G., Matsuoka, Y., and Rebeck, G. W. (2005). F-spondin interaction with the apolipoprotein E receptor ApoEr2 affects processing of amyloid precursor protein.摩尔。细胞杂志。25 (21),9259 - 9268。doi: 10.1128 / mcb.25.21.9259 - 9268.2005
Jakab,。,Schlosser, G., Feijlbrief, M., Welling-Wester, S., Manea, M., Vila-Perello, M., et al. (2009). Synthesis and antibody recognition of cyclic epitope peptides, together with their dimer and conjugated derivatives based on residues 9-22 of herpes simplex virus type 1 glycoprotein D.Bioconjug化学。20 (4),683 - 692。doi: 10.1021 / bc800324g
Karaoglu Hanzatian D。施瓦兹,。,Gizatullin, F., Erickson, J., Deng, K., Villanueva, R., et al. (2018). Brain uptake of multivalent and multi-specific DVD-Ig proteins after systemic administration.马伯10 (5),765 - 777。doi: 10.1080 / 19420862.2018.1465159
Kurokawa, S。,Bellinger, F. P., Hill, K. E., Burk, R. F., and Berry, M. J. (2014). Isoform-specific binding of selenoprotein P to the beta-propeller domain of apolipoprotein E receptor 2 mediates selenium supply.生物。化学。289 (13),9195 - 9207。doi: 10.1074 / jbc.M114.549014
李,b。,Huang, J. S., Jayathilaka, L. P., Lee, J., and Gupta, S. (2016). Antibody production with synthetic peptides.摩尔。生物方法。1474年,第45期。doi: 10.1007 / 978 - 1 - 4939 - 6352 - 2 _2
麦科伊,a . J。,Grosse-Kunstleve, R. W., Adams, P. D., Winn, M. D., Storoni, L. C., and Read, R. J. (2007). Phaser crystallographic software.j:。Crystallogr。40 (4),658 - 674。doi: 10.1107 / S0021889807021206
三角,S。Endo, M。,Mukai, R., Tachibana, K., Umeda, M., Honda, T., et al. (2003). A novel cyclic peptide immunization strategy for preventing HIV-1/AIDS infection and progression.生物。化学。278 (34),32335 - 32343。doi: 10.1074 / jbc.M301209200
Munji r . N。,Soung, A. L., Weiner, G. A., Sohet, F., Semple, B. D., Trivedi, A., et al. (2019). Profiling the mouse brain endothelial transcriptome in health and disease models reveals a core blood-brain barrier dysfunction module.Nat。>。22 (11),1892 - 1902。doi: 10.1038 / s41593 - 019 - 0497 - x
Murshudov g . N。Skubak, P。,Lebedev, A. A., Pannu, N. S., Steiner, R. A., Nicholls, R. A., et al. (2011). REFMAC5 for the refinement of macromolecular crystal structures.Acta Crystallogr。d杂志。Crystallogr。67 (4),355 - 367。doi: 10.1107 / S0907444911001314
Pardridge, w . M。Buciak, j·L。,和Friden, P. M. (1991). Selective transport of an anti-transferrin receptor antibody through the blood-brain barrier在活的有机体内。j .杂志。其他实验。259 (1),66 - 70。
Pardridge, w . M。康,y。,Buciak, J. L., and Yang, J. (1995). Human insulin receptor monoclonal antibody undergoes high affinity binding to human brain capillaries在体外和快速transcytosis穿过血脑屏障在活的有机体内在灵长类动物。制药。Res。12 (6),807 - 816。doi: 10.1023 /: 1016244500596
Saetang, J。,Roongsawang, N., Sangkhathat, S., Voravuthikunchai, S. P., Sangkaew, N., Prompat, N., et al. (2022). Surface cysteine to serine substitutions in IL-18 reduce aggregation and enhance activity.PeerJ10,e13626。doi: 10.7717 / peerj.13626
Simonovic M。Dolmer, K。黄,W。,Strickland, D. K., Volz, K., and Gettins, P. G. (2001). Calcium coordination and pH dependence of the calcium affinity of ligand-binding repeat CR7 from the LRP. Comparison with related domains from the LRP and the LDL receptor.生物化学40 (50),15127 - 15134。doi: 10.1021 / bi015688m
Stanimirovic D。Kemmerich, K。,Haqqani, A. S., Sulea, T., Arbabi-Ghahroudi, M., Massie, B., et al. (2015).胰岛素样生长因子1受体的特殊抗体和使用。专利WO2015131256A1。n . r . c . o .加拿大。加拿大。
Terstappen, g . C。迈耶,a . H。,贝尔,r D。,和Zhang, W. (2021). Strategies for delivering therapeutics across the blood-brain barrier.Nat。启药物。20 (5),362 - 383。doi: 10.1038 / s41573 - 021 - 00139 - y
Valjakka, J。,Hemminki, A., Niemi, S., Soderlund, H., Takkinen, K., and Rouvinen, J. (2002). Crystal structure of an在体外亲和力,specificity-matured anti-testosterone工厂与睾酮在复杂。改进关联变量域内小结构变化的结果。生物。化学。277 (46),44021 - 44027。doi: 10.1074 / jbc.M208392200
Vonrhein C。,Flensburg, C., Keller, P., Sharff, A., Smart, O., Paciorek, W., et al. (2011). Data processing and analysis with the autoPROC toolbox.Acta Crystallogr。d杂志。Crystallogr。67 (4),293 - 302。doi: 10.1107 / S0907444911007773
吴,D。,和Pardridge, w . M。(1999). Blood-brain barrier transport of reduced folic acid.制药。Res。16 (3),415 - 419。doi: 10.1023 /: 1018829920158
Yasui, N。,Nogi, T., and Takagi, J. (2010). Structural basis for specific recognition of reelin by its receptors.结构18 (3),320 - 331。doi: 10.1016 / j.str.2010.01.010
Yu y . J。,Zhang, Y., Kenrick, M., Hoyte, K., Luk, W., Lu, Y., et al. (2011). Boosting brain uptake of a therapeutic antibody by reducing its affinity for a transcytosis target.科学。Transl。地中海。3 (84),84 ra44。doi: 10.1126 / scitranslmed.3002230