Parthenolide诱发快速硫醇氧化导致ferroptosis肝癌细胞
 弗朗西斯卡诉LoBianco1、2,金伯利j . Krager1、2,
弗朗西斯卡诉LoBianco1、2,金伯利j . Krager1、2, 艾丽卡约翰逊
1、2,克里斯托弗·o·古德温1、2,Antino r·艾伦1、2,
艾丽卡约翰逊
1、2,克里斯托弗·o·古德温1、2,Antino r·艾伦1、2, 彼得·a .骗子
2,塞萨尔·m·密友2,迈克尔·j·保华3和
彼得·a .骗子
2,塞萨尔·m·密友2,迈克尔·j·保华3和 Nukhet Aykin-Burns
1、2*
Nukhet Aykin-Burns
1、2*- 1部门的辐射卫生、制药科学系的阿肯色大学医学科学,小石城,AR,美国
- 2阿肯色大学制药科学系的医学科学,小石城,美国基于“增大化现实”技术
- 3阿肯色大学放射学系为医学科学,小石城,基于“增大化现实”技术,美国
肝细胞癌(HCC)是毁灭性的和普遍的疾病。在美国,每年约24500名男性和10000名女性被诊断为肝癌,和超过一半的诊断病人死于这种疾病。到目前为止,传统的治疗没有成功的肝细胞癌患者由于各种潜在的并发症。可怜的存活率和高治疗后复发的发生率之间的差异表明,氧化还原环境周围正常肝和肝癌是有价值的目标,提高治疗效果。Parthenolide (PTL)是一种自然发现治疗具有抗癌和抗炎的特性。PTL可以改变肝癌的抗氧化剂环境通过硫醇修饰使肿瘤细胞敏感升高活性氧(ROS)。调查改变了硫醇之间的联系机制,增加敏感度iron-mediated脂质过氧化作用将允许提高肝癌的治疗。HepG2(人类)和McARH7777(老鼠)肝癌细胞治疗与PTL增加浓度减少细胞生存能力和单独使用效率在体外。PTL增加谷胱甘肽(GSH)氧化获救的谷胱甘肽前体,防治(NAC)。此外,这个海拔硫醇氧化导致线粒体功能障碍的整体提升。阐明如果细胞死亡是通过脂质过氧化作用,使用传感器表示PTL增加脂质氧化脂质过氧化水平后6 h。此外,免疫印迹显示谷胱甘肽过氧化物酶4 (GPx4)蛋白质含量降低与PTL暗示治疗后细胞暴露在PTL后无法阻止脂质过氧化作用。海拔在脂质过氧化作用将导致细胞死亡的一种形式被称为ferroptosis。进一步建立ferroptosis作为肝癌死亡的关键机制在体外,添加ferrostatin-1结合PTL演示了一个殖民地生存分析的部分恢复。这项研究表明,PTL可以诱导肿瘤细胞死亡在细胞内氧化、海拔ferroptosis离开细胞敏感。
1介绍
肝细胞癌(HCC)是主要的癌症肝细胞和估计导致2025 < 100万新病例(Llovet et al ., 2021)。有各种与肝癌相关的风险因素,包括肝硬化、病毒性肝炎、酒精、肥胖、和非酒精性脂肪肝病(NAFLD) (黄et al ., 2021)。一个包罗万象的机制与这些危险因素是一个损伤肝脏正常的薄壁组织,导致进行性炎症和再生,导致肝癌的最终发展(马雷罗,2020)。5年相对生存患者诊断为肝癌患者只有18%,减少局部肿瘤(西格尔et al ., 2019)。这个可怜的生存的一个因素是,HCC诊断通常是在其课程设置内的慢性肝脏疾病。典型的晚期诊断hcc诊断后导致中等生还的6 - 12个月(调查人员,1998)。虽然有各种治疗肝癌的选择,并不是所有的选项可用病人由于肝癌的严重程度和健康的肝脏出现(El-Serag et al ., 2008)。肝细胞癌是由于高表达的耐多药耐药肿瘤蛋白质和高活性氧(ROS) (Ng et al ., 2000;Ogunwobi et al ., 2019)。总的来说,需要新的疗法,可以负担得起的,有效的,并发表对肝癌局部区域。
Parthenolide (PTL)是一种自然产生的倍半萜烯内酯是从植物菊科植物(Tanacetum光)。PTL一直用于治疗关节炎、虫咬、感染、头痛、感冒(1999年查韦斯和查韦斯;Jain和Kulkarni, 1999年)。然而,最近,PTL被发现有抗癌和pro-apoptotic能力(温家宝et al ., 2002)。先前的研究属性PTL细胞毒性活动与硫醇内酯基的反应,如蛋白质,含有半胱氨酸残基或含巯基的团体(sh) (赫兹,1977;Zhang et al ., 2004)。谷胱甘肽(GSH)是一个重要的硫醇和氧化还原衬底对于细胞内的活性氧的解毒和被氧化的速率,控制挤压、硫醇前兆的吸收,合成(藤井裕久et al ., 2011)。在结直肠癌细胞,损耗线粒体膜上的巯基共同损害线粒体膜的完整性造成氧化应激的恶性循环癌症细胞死亡(Zhang et al ., 2004)。先前的研究表明,PTL激活NADPH氧化酶类,增加了ROS在前列腺癌细胞,抑制抗氧化剂产生氧化应激升高(太阳et al ., 2010)。在这项研究中,我们证明PTL不仅导致谷胱甘肽水平降低和海拔的肿瘤细胞内ROS,但这谷胱甘肽的耗竭导致下游的细胞死亡形式称为ferroptosis。
Ferroptosis是调节细胞死亡的一种形式定义的缺失或不足selenoperoxidase活动,谷胱甘肽过氧化物酶4 (GPx4),导致增加活性氧的水平和细胞死亡通过脂质氧化膜(Yu et al ., 2016;Lei et al ., 2019;谅解备忘录et al ., 2019)。Ferroptosis细胞阻碍进口半胱氨酸通过Xc-transport系统合成谷胱甘肽GPx4活动。细胞外半胱氨酸是进口的通过半胱氨酸/谷氨酸转运体。然后利用半胱氨酸形成谷胱甘肽,GPx4的衬底。GPx4转换PUFA-OOH PUFA-OH减少的数量可能在细胞内活性氧。PTL与谷胱甘肽反应预防足够GPx4功能,导致氧化损伤和最终通过ferroptosis细胞死亡。电流通过ferroptosis包括erastin药物用于治疗癌症,索拉非尼和柳氮磺胺吡啶抑制Xc-system通过抑制半胱氨酸运输从而阻止谷胱甘肽合成和GPx4函数(Yu et al ., 2016;谅解备忘录et al ., 2019)。
谷胱甘肽是重要的基质GPx4,降低谷胱甘肽细胞导致死亡的一种形式形态相同GPx4的死亡引起的损失(迪克森et al ., 2012)。通常在正常细胞,抗氧化机制有效地防止脂质过氧化GPx4发起的活动,它使用谷胱甘肽(斯科特,1997)。有趣的是,虽然癌症通常有高水平的ROS和氧化还原代谢特异表达,ferroptosis不经常发生在肿瘤发生。此外,触发ferroptosis诱发therapy-resistant癌症死亡由于癌细胞的生存依靠GPx4 (Hangauer et al ., 2017)。原因GPx4依赖性是癌细胞依赖于铁水平比正常细胞的增长,新陈代谢,和转移导致铁作为催化剂可以迅速脂质过氧化脂质膜是否不受GPx4保护(刘et al ., 2018)。最近,PTL与ferroptosis在三阴乳腺癌(TNBC)细胞ROS高程和GPx4泛素化(丁et al ., 2021)。然而,没有研究关注ferroptosis死亡作为一种机制在HCC倍半萜烯内酯。这项研究表明,PTL诱发ferroptosis谷胱甘肽耗竭后肝癌,快速氧化细胞内线粒体硫醇,线粒体功能障碍,抑制GPx4。
2材料和方法
2.1细胞和培养条件
细胞的选择在体外实验包括人类肝细胞癌(HepG2)和莫里斯鼠肝癌(McA-RH7777)从美国获得细胞类型文化集合(写明ATCC马纳萨斯,弗吉尼亚州,美国)。细胞培养在各自媒体最低基本媒体(MEM)和杜尔贝科的修改鹰介质(DMEM)补充10%胎牛血清的边后卫和100 U /毫升青霉素和链霉素100μg /毫升(P / S)。细胞被保持在5%的股份有限公司2调湿大气在37°C。收集细胞实验用0.25%胰蛋白酶和伯爵夫人用台盼蓝染色计数细胞计数器来确认90%或更好的生存能力。PTL(σ,伯灵顿,妈,美国)获得和可溶性二甲亚砜(DMSO) 25毫米的浓度,然后稀释浓度在PBS。车辆的最终浓度DMSO溶液治疗组为0.05%。
2.2隔离小鼠原发性肝细胞
原发性肝细胞分离得到C57BL / 6小鼠通过中空的下腔静脉灌注肝用含氧钙免费灌注缓冲区在37°C(汉克斯平衡盐溶液,5.8毫米消息灵通的,4.5毫米NaHCO3)。肝组织灌注后,含氧消化与5毫米CaCl缓冲区(灌注缓冲区2、15毫克胶原酶和25 mg胰蛋白酶抑制剂)在37°C添加到肝脏开始消化。肝脏被去除肝脏进一步消化和把它放在培养皿中un-supplemented F-12媒体。细胞被旋转在4°C 3分钟800转两次。细胞在25毫升resuspended un-supplemented F-12媒体和25毫升的90% PerColl和离心机在4°C 8分钟140 g移除碎片和死细胞。细胞被resuspended补充(10%的边后卫,100 U /毫升青霉素和链霉素100μg /毫升)染料自由RPMI 1640媒体和离心机在800转3分钟去除残余PerColl。颗粒是resuspended RPMI 1640年完成媒体和台盼蓝染色计数。只有90%或以上可行性的集合被用于实验。收集后10000细胞/涂镀在胶原IV 96板进行进一步的实验。
2.3 MTT试验
5-dimethylthiazol-2-yl 3 - (4) 2, 5-diphenyltetrazolium溴化(MTT)试验是用来评估所需的剂量的PTL进一步细胞培养实验。HepG2、mcarh - 7777和孤立的原发性肝细胞被镀96板在3000 - 10000细胞。治疗组在一式四份完成越来越多的PTL以及车辆控制(0.05% DMSO),媒体控制和有毒的10% DMSO溶液。板暴露在治疗24 h。另外一个盘HepG2细胞治疗10毫米NAC PTL治疗前24小时。细胞孵化与MTT(500μg /毫升最终浓度井)在37°C 4 h。包含媒体被MTT, DMSO溶液添加到井和盘子被放置在一个轨道瓶5分钟后吸光度测量在490 nm使用BioTek板读者。MTT还原控制(%)计算每组和放置在对数尺度来确定每个细胞株的IC50剂量和IC90剂量为未来的实验通过非线性回归分析。
2.4单独生存分析
HepG2和mcarh - 7777细胞治疗的500海里ferrostatin-1 PTL IC50,治疗前24小时IC90 3 h和6 h。治疗后细胞使胰蛋白酶化,re-plated单独使用不同密度,留给10 - 14天37°C孵化器。殖民地与70%乙醇固定,与考马斯亮蓝染色剂染色(准备在50%甲醇,5%的醋酸,45%的H2O)并使用显微镜计数。殖民地被定义为每殖民地和50细胞百分比决定生存。
2.5硫醇氧化状态的细胞
HepG2 mcarh - 7777和50 - 100处理莫伊胞质或线粒体roGFP腺病毒后以前公布的协议(阿利耶夫et al ., 2020)。细胞被对待IC50, PTL IC90剂量。改变氧化和减少之间的荧光波长频率测量和量化使用荧光显微镜成像可视化和代表。量化后的细胞时间点30分钟,1 h, 2 h, 4 h, 6小时,24小时由流式细胞术(BD LSRFortessa)封闭的渠道激励405海里/发射525海里(510年灿烂的紫[BV510])和励磁488海里/发射525海里(异硫氰酸荧光素(FITC)的协助下解决分子成像的核心设备。每个细胞的比例从激发荧光在405和488海里,抵消了指标的数量和绝对光学灵敏度,表明氧化发生。
2.6脂质过氧化反应的评估
HepG2细胞治疗在IC50 IC90 PTL 30分钟,3 h, 6 h时间点。10µM图像,脂质过氧化反应传感器(表达载体,沃尔瑟姆,妈,美国)添加到细胞和孵化为30分钟,细胞被洗PBS。氧化和减少的比率是488年由激发/发射510 nm (TexasRed)和励磁581 nm /发射591海里(FITC)流式细胞术(BD LSRFortessa)协助解决分子成像的核心设备。
2.7总和氧化谷胱甘肽测定
1×106HepG2细胞治疗PTL IC50一式三份,IC90剂量3 h, 6 h和24小时在两个独立的日子。额外的细胞治疗10毫米NAC治疗前24小时PTL一式三份同样的时间点。细胞被收集和稀释在1.34毫米DETAPAC (diethylenetriaminepenta-acetic酸)缓冲区在冰上。细胞细胞溶解在5%磺基水杨酸和离心机。5、5′-dithiobis-2-nitrobenzoic酸回收试验进行测量总谷胱甘肽和GSSG水平(nmol /毫克蛋白)使用UV / Vis分光光度计(珀金埃尔默格里菲斯,1980)。数据归一化蛋白质含量由洛瑞的蛋白质测定(洛瑞et al ., 1951)。
2.8测量的线粒体功能
HepG2细胞被镀10000每口井Celltak涂板和媒体改为无缓冲的DMEM non-CO包含4毫米谷氨酸和孵化2孵化器在37°C 1 h。IC50, IC90剂量注入和四个基线测量之前获得线粒体抑制剂或解偶联剂的注入。读数后拍摄顺序添加寡霉素(10µM), FCCP(5µM),和鱼藤酮/抗霉素A(10µM),和测量24 h。耗氧速率(OCR)以37°C使用XF96细胞外流量分析仪(安捷伦科技)。OCR被海马XF96计算软件和代表平均20-32测量在不同的日子里(Ferrick et al ., 2008)。
2.9免疫印迹
HepG2细胞被镀1×106和接受IC50 IC90剂量1 h, 3 h, 6 h和24小时,细胞被收集。蛋白质浓度进行了分析使用皮尔斯BCA蛋白质化验设备和标准曲线(23225年,热费希尔,美国)。蛋白质样品被稀释在1 x Laemmli样品缓冲5% 2β巯基乙醇(2-BME)和均匀加载与25µg蛋白质/巷5 - 14%梯度Mini-PROTEAN TGX凝胶(Bio-Rad 456 - 1034或456 - 1084年,美国)。凝胶是运行60分钟90 mV和转移到硝基吸纸和运行60分钟100 mV在4°C。1 h墨迹被封锁的奥德赛阻断缓冲区(NE LI-COR,林肯,美国)然后孵化1:1000目标抗体的稀释GPx4一夜之间(ab125066 Abcam,妈,美国)。加载控制抗体lamin-A / C孵化3 h(4777年,细胞信号,妈,美国)。洗后,这些墨迹被孵化的1:10,000稀释LI-COR反鼠标/ anti-rabbit荧光抗体40分钟。屁股被存储在1 x PBS。成像是由使用一个可视化奥德赛Fc成像系统(NE LI-COR,林肯,美国)。墨迹被暴露在700年和800年的λ2分钟每个通道。分析微与LI-COR软件完成。
2.10统计分析
9.1.2 GraphPad棱镜版本被用来执行统计分析。数据表示为±SEM。确定统计学意义p< 0.05,表示为*p< 0.05,* *p< 0.01,* * *p< 0.005。IC50值是由非线性回归分析从块MTT还原控制(%)与LogC (M)的药物浓度和一个未配对的双尾t以及一式三份措施之间进行比较的IC50 PTL和南汽。意义决定使用单向方差分析和图基硫醇氧化后分析,谷胱甘肽(GSSG测量,脂质过氧化反应,GPx4蛋白质水平和耗氧率。双向方差分析与Sidak多个用于事后比较分析ferrostatin-1救援单独测定PTL单独分析。
3的结果
3.1 PTL是肝癌细胞的细胞毒性
HepG2和mcarh - 7777细胞暴露于浓度增加PTL为未来实验确定合适的浓度。DMSO溶液,用作车辆不改变细胞毒性或任何其他参数我们测量在整个研究(补充数据S1-S3)。MTT还原控制(%)稳步降低PTL的浓度增加。IC90剂量被选为三倍的IC50实验揭示细胞反应的差异在24 h。图1 bHepG2 IC50:18μM IC90: 54µM和图1 cMcA-RH7777 IC50:13µM IC90: 40µM。人类和鼠肝癌行之间重要的是,IC50剂量分别为18μM和13µM中是相似的。此外,PTL暴露在小鼠原发性肝细胞分离建议PTL正常肝组织的毒性还是比癌症细胞系的IC50剂量的8.8毫米(图1一个)。
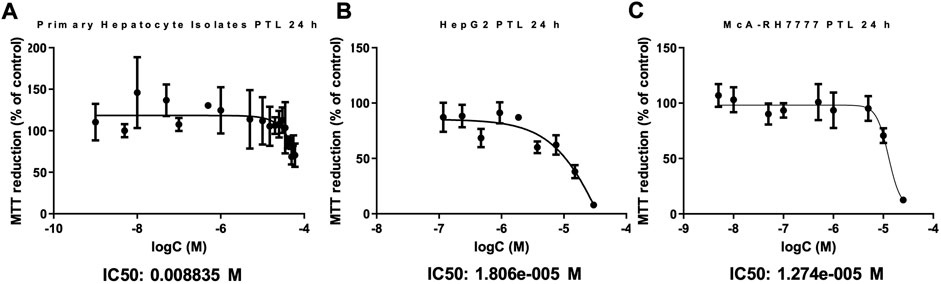
图1。。PTL大体上是主要的小鼠肝细胞的毒性较小(一)比HepG2(B)和McARH7777(C)细胞。MTT试验被用来估计主要小鼠肝细胞的细胞生存能力(一)HepG2(人类)(B),McARH7777(老鼠)(C)细胞。板块与浓度增加孵化PTL为24小时一式四份(n = 12)在三个不同的实验。非线性回归分析确定IC50和IC90 PTL为每个。(一)IC50: 8.8毫米,IC90: 26毫米。(B)IC50: 18µM IC90: 54µM。(C)IC50: 13µM IC90: 40µM。
3.2 PTL迅速氧化细胞内硫醇
硫醇氧化被认为是一个重要的机制PTL细胞内的快速交互。RoGFP是一个传感器,可以送到细胞adenovirally评估活细胞硫醇氧化发生(阿利耶夫et al ., 2020)。图2演示PTL的能力迅速氧化HepG2细胞内胞质硫醇,PTL以来持续超过24小时。在之前的研究发现导致线粒体的改变(Zhang et al ., 2004),我们使用了一个线粒体roGFP传感器评估线粒体硫醇氧化,发现在IC90剂量有几乎三倍增加线粒体氧化硫醇后24小时(图2 c)。PTL导致持续的氧化线粒体硫醇可能导致线粒体功能障碍的过程。

图2。胞质PTL诱发海拔(B)和线粒体(C)硫醇氧化。(一)代表由荧光显微镜成像获得的。(B)Adenovirally表示比率计氧化状态标记RoGFP用流式细胞仪测量405 nm / em。525 nm和488 nm / em。525海里显示海拔在整个细胞内氧化,以应对PTL早在1 h后处理。(C)PTL导致海拔线粒体硫醇氧化后24 h。(n = 3)。错误代表±1 SEM。意义由单向方差分析(#p< 0.05)与图基后分析比较控制IC50(18µM)和IC90(54µM) *p< 0.05,* *p< 0.01,* * *p< 0.001。
3.3主要的硫醇,谷胱甘肽(GSH)是由PTL氧化,将减少和防治作用的氧化
一个主要的硫醇在细胞内谷胱甘肽。谷胱甘肽具有宝贵的作用的基本细胞的氧化还原体内平衡,因为它可以不断地氧化和减少细胞内的酶清除活性氧(ROS) (福尔曼et al ., 2009)。有趣的是,图3 a - c演示了一个区别在细胞反应PTL IC50剂量和IC90剂量之间的关系。当细胞暴露在IC50剂量24 h,细胞的反应是提高水平的减少谷胱甘肽(图3一),减少氧化谷胱甘肽也称为GSSG (图3 b, C)。细胞能够克服快速氧化谷胱甘肽通过补偿机制然而当细胞接受IC90剂量超过24小时,细胞无法克服这个增加谷胱甘肽氧化表示GSSG谷胱甘肽和海拔的降低。

图3。PTL增加谷胱甘肽(GSH)在HepG2细胞氧化(B, C)。细胞试图克服提升减少谷胱甘肽氧化的IC50 PTL(18µM)剂量(一)。然而,IC90(54µM)剂量预防补偿性反应氧化增加,导致细胞死亡(一)。10毫米NAC获救的谷胱甘肽水平细胞治疗IC90剂量(D)和减少氧化谷胱甘肽的水平(E, F)。(两者)HepG2细胞治疗PTL (IC50或IC90)每2天分别计算一式三份(n = 6)。谷胱甘肽(一),GSSG(B)和% GSSG(C)规范化的蛋白质含量。错误代表±1 SEM。意义由单向方差分析(#p< 0.05)与图基后分析比较控制IC50(18µM)和IC90(54µM) *p< 0.05,* *p< 0.01,* * *p< 0.001。(D-F)HepG2细胞治疗10毫米NAC治疗前24小时IC90剂量的2天分别为每个计算PTL一式三份(n = 6)。谷胱甘肽(一),GSSG(B)和% GSSG(C)规范化的蛋白质含量。错误代表±1 SEM。之间的意义由单向方差分析与图基后分析IC90(54µM)和NAC-IC90 *p< 0.05,* *p< 0.01,* * *p< 0.001。
当一个谷胱甘肽前体(NAC)称为防治增加了24小时PTL治疗之前,这些细胞能够提高谷胱甘肽水平,以应对氧化发生的数量从PTL (图3 d)。此外,图3 e, F显示更少的谷胱甘肽氧化成GSSG后24 h (PTL)。有趣的是,南汽的细胞治疗IC90剂量表现类似于IC50细胞治疗图3 a - c通过细胞内的谷胱甘肽存在。图3 d-f表明,NAC能有效地救助谷胱甘肽的水平和硫醇氧化但不能完全防止氧化出席IC90剂量。
3.4南京营救PTL的细胞毒性
当我们在图3,NAC可以保护肝细胞谷胱甘肽氧化。图4演示了硫醇氧化导致的预防保护PTL在人类细胞毒性(图4一)和大鼠(图4 b)肝癌细胞。这10毫米的NAC可以增加细胞的能力,减少肝癌甚至在最高剂量的PTL和显著增加所需的IC50剂量达到相同的毒性。对HepG2细胞PTL IC50剂量为18µM但的NAC 48µM IC50剂量(图4一)。McA-RH7777细胞PTL IC50剂量为13µM但的NAC IC50剂量增加到21µM (图4 b)。

图4。10毫米NAC逆转减少MTT还原HepG2的控制(%)(一)和McARH7777(B)细胞PTL浓度增加。NAC PTL IC50剂量增加到48µM HepG2(一)和21µM McA-RH7777。MTT试验被用来估计HepG2细胞生存能力(一)和McARH7777(B)。板块与10毫米孵化NAC治疗前24小时浓度增加PTL一式四份24 h 3独立实验。非线性回归分析来确定的IC50剂量PTL为每个(n = 12)。错误代表±1 SEM。之间的意义由未配对的双尾学习一式三份的措施。(一)IC50(18µM)和NAC-IC50 HepG2(48µM)。(B)IC50(13µM)和NAC-IC50 McA-RH7777 * *(µM 21日)p< 0.01,* * *p< 0.001。
3.5 PTL增加脂质过氧化,减少GPx4出现在肝癌细胞的数量
谷胱甘肽与GPx4发挥着重要作用,清除细胞内的脂质氧化可能发生的。因为我们发现硫醇的快速氧化,导致减少HepG2细胞内谷胱甘肽,我们评估了脂质过氧化的礼物。图5表明脂质过氧化增加6 h后接触的IC50和IC90剂量HepG2细胞。高脂质过氧化作用是一个值得注意的机制,启动细胞死亡的形式被称为ferroptosis。类似最近发现在三阴性乳腺癌细胞(丁et al ., 2021),图6表明在HepG2细胞,GPx4蛋白质含量明显下降,但只有在IC90剂量后24 h。有趣的是,复苏的IC50剂量允许GPx4水平可能占一些两剂之间的反应的差异。

图5。PTL HepG2细胞诱导脂质过氧化反应。脂质过氧化作用来衡量图像传感器和流式细胞术显示海拔与PTL治疗后脂质过氧化作用。氧化和减少的比率由488 nm / em。510 nm和581 nm / em。591海里。叠化决心相比之下汽车控制(n = 3)。错误代表±1 SEM。意义是由单向方差分析与图基后分析。IC50(18µM)和IC90(54µM) *p< 0.05,* *p< 0.01。

图6。HepG2细胞治疗后PTL证明减少GPx4当暴露在IC90 PTL(54µM)剂量。细胞暴露在IC50(18µM)剂量似乎增加GPx4目前的水平。成像(一)和分析(B)获得使用Li-Cor成像软件。污点是规范化的加载控制。叠化是由微车辆控制的比较(n = 3)。错误代表±1 SEM。意义由单向方差分析(# p < 0.0001)与图基的文章分析比较控制IC50(18µM)和IC90(54µM) *p< 0.05。显著区别IC90 1 h和IC90 24 h * *p< 0.01。#p< 0.0001 IC50, IC90代表一个重要的区别。
3.6抑制ferroptosis部分保护细胞免受PTL
示范高程的脂质过氧化作用图5的减少GPx4目前在肝癌细胞(图6)表示ferroptosis可能是细胞死亡的机制。确认ferroptosis发生接触PTL之后,我们添加了ferrostatin-1 (ferroptosis抑制剂)与PTL细胞治疗之前。细胞生存分析显示部分人类(6 h后的救援图7)和大鼠(图7 b)肝癌细胞。图7表明ferroptosis是细胞死亡的机制但不是唯一机制自细胞并不完全由ferroptosis抑制获救。

图7。HepG2(一)和McA-RH7777(B)细胞获救ferroptosis抑制剂(ferrostatin-1)单独生存分析。细胞被镀在不同clonigenic密度与500 nM Ferrostatin-1 PTL治疗或治疗后,PTL治疗前24小时(一)IC50(18µM), IC90µM(54),或(B)IC50(13µM)和IC90(40µM) (n = 6 - 12单独使用盘子镀3单独治疗菜)。错误代表±1 SEM。意义由单向方差分析与图基后分析*p< 0.05。
3.7 PTL导致线粒体功能障碍
由于线粒体硫醇氧化24 h后暴露在PTL升高(图2 c),我们想要评估这氧化干扰线粒体功能和呼吸。使用海马通量分析仪,图8表明整体基础呼吸(图8)后迅速下降1 h。另外,有一个减少线粒体耦合效率(图8 f)和ATP与线粒体呼吸(图8 b)表明PTL暴露线粒体是不再有效地生成ATP。此外,电子传递链泄漏更多的质子(图8 c)进一步支持这样的结论:线粒体功能异常。总的来说,PTL线粒体硫醇氧化提供了一个早期的主要破坏线粒体活动会导致线粒体呼吸效率低下的恶性循环,生成更多的氧化应激和细胞死亡。

图8。PTL HepG2细胞导致线粒体功能障碍所示基底减少呼吸(一),ATP-linked呼吸(B),海拔在质子泄漏(C),减少最大的呼吸(D)和储备呼吸能力(E),%降低耦合(F)。HepG2细胞治疗的IC50(18µM)或IC90(54µM)剂量的PTL线粒体解偶联剂或抑制剂注入之前确定耗氧速率(OCR)在1 h和3 h (n = 3)错误代表±1 SEM。意义由单向方差分析与图基后分析*p< 0.05,* *p< 0.01,* * *p< 0.001,* * * *p< 0.0001。
4讨论
PTL是一种天然提取物镇痛,抗菌、抗炎、抗癌机制,依靠广泛的细胞信号(Zhang et al ., 2004)。PTL体内的行动似乎没有一个机制,但各种相互作用的分子靶点等核因子kappa-light链(NF-κB),激活NADPH氧化酶类,硫醇反应和诱导细胞凋亡(Sobota et al ., 2000;Pozarowski et al ., 2003;古兹曼et al ., 2005;太阳et al ., 2010)。先前的研究已经表明,PTL肝癌细胞内抗癌活动将暂停在G2 / M期细胞周期进展通过G0-G1 p53激活并逮捕了细胞周期蛋白D1的减少表达式(温家宝et al ., 2002;Ralstin et al ., 2006)。同样,PTL HepG2细胞诱导凋亡通过减少bcl - 2表达的抗凋亡和高架pro-apoptotic蛋白质,导致最终激活caspase-9和caspase-3 (太阳et al ., 2014)。然而,细胞凋亡并不是唯一的细胞死亡机制以应对PTL,自噬也发生在PTL HepG2细胞内暴露,胰腺癌细胞,骨肉瘤细胞,宫颈癌(太阳et al ., 2014;D 'anneo et al ., 2013;Jeyamohan et al ., 2016;刘et al ., 2017)。我们目前的研究证实,细胞凋亡并不是唯一的细胞死亡机制发生在PTL ferroptosis发挥了重要作用在肝癌细胞的细胞毒性效应。
高氧化应激会导致氧化损伤,创建一个前馈串级的增加产生活性氧和导致细胞死亡(Berlett Stadtman, 1997)。先前的研究认为PTL活动快速反应的内酯基硫醇(赫兹,1977)。目前的研究发现,PTL迅速耗尽细胞内谷胱甘肽等硫醇通过增加迅速氧化和氧化线粒体硫醇。这些发现在结肠癌细胞中发现的相类似的,在消耗线粒体膜上的巯基共同导致损伤线粒体膜的完整性,导致氧化应激的恶性循环癌症细胞死亡(Zhang et al ., 2004)。我们发现在肝癌细胞中,线粒体的海拔硫醇氧化导致线粒体呼吸和效率的迅速恶化后1小时。添加谷胱甘肽前体,有趣的是,南汽,24小时之前能够移植与IC90 HepG2细胞谷胱甘肽水平,治疗剂量最初枯竭的谷胱甘肽水平得以补偿,更有效地应对PTL平衡谷胱甘肽的水平。此外,NAC能拯救PTL肝癌细胞的细胞毒性试验,表明PTL硫醇氧化是一个重要的机制在肝癌细胞毒性。这项研究表明从PTL硫醇氧化发生很快,导致早期线粒体功能障碍和氧化应激。
下游影响氧化池增加细胞内谷胱甘肽和快速氧化硫醇会离开细胞容易受到其他氧化反应的氧化脂质(Ursini et al ., 1989)。使用图像,脂质过氧化反应传感器,我们演示了脂质过氧化增加后6 h (PTL在肝癌细胞。这种高度的脂质过氧化作用使我们考虑后续ferroptosis的细胞机制。Ferroptosis是细胞死亡的一种形式,从失踪或GPx4活动不足,导致脂质过氧化增加(Lei et al ., 2019)。这是第一次研究证明ferroptosis肝癌细胞中细胞死亡的机制。我们添加ferrostatin-1能够部分恢复细胞表明ferroptosis发生;然而,随着PTL的其他细胞死亡机制,如细胞凋亡和自噬,我们无法完全拯救细胞。ferroptosis仍然是一个独特的、不可逆转的细胞死亡机制,引发ferroptosis将导致therapy-resistant癌症死亡由于癌细胞的生存依靠GPx4 (Hangauer et al ., 2017)。我们证明PTL减少GPx4 HepG2细胞内的蛋白质含量,类似于发现了什么丁et al。(2021),提出了结合ferroptosis和细胞凋亡机制三阴性乳腺癌细胞的细胞死亡。失去GPx4可能表明为什么这海拔的脂质过氧化和ferroptosis发生在后来的时间点。
重要的是,PTL明显微分影响肿瘤细胞与正常细胞相比。的一项研究温家宝et al。(2002)报道,PTL增加总谷胱甘肽non-tumoral肝细胞而引起癌症肝细胞谷胱甘肽的耗竭。此外,这种高度的谷胱甘肽还发现在正常前列腺细胞辐射防护(太阳et al ., 2010;徐et al ., 2013)。同样地,我们发现PTL孤立主小鼠肝细胞的毒性远远比肝癌细胞株表明使用细胞毒性剂量的升高PTL治疗肝癌不会正常肝实质损害。然而,使用PTL的一个重大缺点是其低生物利用度主要是亲脂性的性质(赵et al ., 2016)。这个独特的特征PTL HCC患者可能受益,大部分病人都没有资格获得治疗系统治疗和需要独特的选项如trans-arterial化疗栓塞术(TACE)。别说话直接允许交付治疗治疗肿瘤以最小的副作用通过避免全身新陈代谢(de Baere et al ., 2016;鲜明的et al ., 2020)。别说话会导致60%的反应率持续6个月或更长时间(Llovet Bruix, 2003;Zhang et al ., 2018)。然而,目前,TACE-treated肿瘤往往re-vascularize,导致复发率增加(公园et al ., 2013)。别说话最常用药物阿霉素;然而,大约有50%的肿瘤用阿霉素治疗没有反应,只有30%显示一个完整的响应(林et al ., 2010)。因此,有必要考虑替代选项,确保治疗肝癌患者是有效的。根据这项研究,我们相信PTL可以用作治疗肝动脉化疗栓塞剂,并计划进一步研究在活的有机体内机制和响应PTL交付局部区域在McA-RH777原位肿瘤模型。
数据可用性声明
最初的贡献提出了研究中都包含在这篇文章/补充材料,进一步的调查可以针对相应的作者。
作者的贡献
NA-B、CC和MB导致概念和设计的研究。FL、KK EJ, CG进行实验。FL进行了统计分析。FL写了初稿的手稿。NA-B, CC、AA和KK编辑部分的手稿。所有作者导致修订手稿、阅读和批准提交的版本。
资金
这项工作是在NIH /赠款支持美国P20GM109005 (KK和NA-B);NIH /美国国家T32 GM106999 (FL), NIH / NHLBI R25HL108825 (EJ) UAMS AWD53956绝笔(NA-B);UAMS三飞行员FY19 (MB);NIH / NCI R01CA258673 (AA, FL, NA-B)。投资者没有参与研究设计、数据收集和分析,决定发表,或准备的手稿。
的利益冲突
作者声明,这项研究是在没有进行任何商业或财务关系可能被视为一个潜在的利益冲突。
出版商的注意
本文表达的所有索赔仅代表作者,不一定代表的附属组织,或出版商、编辑和审稿人。任何产品,可以评估在这篇文章中,或声称,可能是由其制造商,不保证或认可的出版商。
补充材料
本文的补充材料在网上可以找到:https://www.雷竞技rebatfrontiersin.org/articles/10.3389/ftox.2022.936149/full补充材料
引用
阿利耶夫,a . T。,Lobianco, F., Krager, K. J., and Aykin-Burns, N. (2020). Assessment of cellular oxidation using a subcellular compartment-specific redox-sensitive green fluorescent protein.j .粘度实验。,e61229。doi: 10.3791/61229
Berlett, b S。,和Stadtman, E. R. (1997). Protein oxidation in aging, disease, and oxidative stress.生物。化学。272年,20313 - 20316。doi: 10.1074 / jbc.272.33.20313
D 'Anneo, A。,Carlisi, D., Lauricella, M., Puleio, R., Martinez, R., di Bella, S., et al. (2013). Parthenolide generates reactive oxygen species and autophagy in MDA-MB231 cells. A soluble parthenolide analogue inhibits tumour growth and metastasis in a xenograft model of breast cancer.细胞。死亡说。4,e891。doi: 10.1038 / cddis.2013.415
de Baere T。时候,Y。,Lencioni, R., Geschwind, J. F., Rilling, W., Salem, R., et al. (2016). Treatment of liver tumors with lipiodol TACE: Technical recommendations from experts opinion.Cardiovasc。间歇雨刷。Radiol。39岁,334 - 343。doi: 10.1007 / s00270 - 015 - 1208 - y
叮,Y。,Chen, X., Liu, C., Ge, W., Wang, Q., Hao, X., et al. (2021). Identification of a small molecule as inducer of ferroptosis and apoptosis through ubiquitination of GPX4 in triple negative breast cancer cells.j .内科杂志。肿瘤防治杂志。14日,19。
迪克逊,s . J。,Lemberg, K. M., Lamprecht, M. R., Skouta, R., Zaitsev, E. M., Gleason, C. E., et al. (2012). Ferroptosis: An iron-dependent form of nonapoptotic cell death.细胞。149年,1060 - 1072。doi: 10.1016 / j.cell.2012.03.042
el-Serag, h . B。,Marrero, J. A., Rudolph, L., and Reddy, K. R. (2008). Diagnosis and treatment of hepatocellular carcinoma.胃肠病学134年,1752 - 1763。doi: 10.1053 / j.gastro.2008.02.090
Ferrick, d . A。尼尔森,。,和Beeson, C. (2008). Advances in measuring cellular bioenergetics using extracellular flux.药物。今天13日,268 - 274。doi: 10.1016 / j.drudis.2007.12.008
福尔曼,h·J。张,H。,和Rinna, A. (2009). Glutathione: Overview of its protective roles, measurement, and biosynthesis.摩尔。Asp。地中海。1 - 12。doi: 10.1016 / j.mam.2008.08.006
藤井裕久,J。,Ito, J.-I., Zhang, X., and Kurahashi, T. (2011). Unveiling the roles of the glutathione redox system在活的有机体内通过分析转基因老鼠。j .中国。物化学。减轻。49岁,70 - 78。doi: 10.3164 / jcbn.10 - 138 sr
格里菲斯,o . w . (1980)。二硫化谷胱甘肽,谷胱甘肽测定使用谷胱甘肽还原酶和2-vinylpyridine。肛交。物化学。106年,207 - 212。0003 - 2697 . doi: 10.1016 / (80) 90139 - 6
古兹曼,m . L。,Rossi, R. M., Karnischky, L., Li, X., Peterson, D. R., Howard, D. S., et al. (2005). The sesquiterpene lactone parthenolide induces apoptosis of human acute myelogenous leukemia stem and progenitor cells.血105年,4163 - 4169。doi: 10.1182 /血液- 2004 - 10 - 4135
Hangauer, m . J。,Viswanathan, V. S., Ryan, M. J., Bole, D., Eaton, J. K., Matov, A., et al. (2017). Drug-tolerant persister cancer cells are vulnerable to GPX4 inhibition.自然551年,247 - 250。doi: 10.1038 / nature24297
黄,d Q。,el-Serag, h . B。,和Loomba, R. (2021). Global epidemiology of NAFLD-related HCC: Trends, predictions, risk factors and prevention.Nat,启杂志。乙醇。18日,223 - 238。doi: 10.1038 / s41575 - 020 - 00381 - 6
调查员,c . o . t . l . i . p . (1998)。一个新的肝细胞癌预后系统:回顾性研究435例:肝癌意大利项目(夹)调查人员。肝脏病学28日,751 - 755。doi: 10.1002 / hep.510280322
耆那教徒:K。,和Kulkarni, S. K. (1999). Antinociceptive and anti-inflammatory effects of Tanacetum parthenium L. extract in mice and rats.j . Ethnopharmacol。68年,251 - 259。doi: 10.1016 / s0378 - 8741 (99) 00115 - 4
Jeyamohan, S。,Moorthy, R. K., Kannan, M. K., and Arockiam, A. J. (2016). Parthenolide induces apoptosis and autophagy through the suppression of PI3K/Akt signaling pathway in cervical cancer.Biotechnol。列托人。38岁,1251 - 1260。doi: 10.1007 / s10529 - 016 - 2102 - 7
林,J。,Malagari, K., Vogl, T., Pilleul, F., Denys, A., Watkinson, A., et al. (2010). Prospective randomized study of doxorubicin-eluting-bead embolization in the treatment of hepatocellular carcinoma: Results of the PRECISION V study.Cardiovasc。间歇雨刷。Radiol。33岁的41-52。doi: 10.1007 / s00270 - 009 - 9711 - 7
Lei, P。,Bai, T., and Sun, Y. (2019). Mechanisms of ferroptosis and relations with regulated cell death: A review.前面。杂志。10日,139年。doi: 10.3389 / fphys.2019.00139
刘,H。,Schreiber, S. L., and Stockwell, B. R. (2018). Targeting dependency on the GPX4 lipid peroxide repair pathway for cancer therapy.生物化学57岁,2059 - 2060。doi: 10.1021 / acs.biochem.8b00307
刘,W。,Wang, X., Sun, J., Yang, Y., Li, W., and Song, J. (2017). Parthenolide suppresses pancreatic cell growth by autophagy-mediated apoptosis.Onco。目标。其他。10日,453 - 461。doi: 10.2147 / OTT.S117250
Llovet, j . M。,和Bruix, J. (2003). Systematic review of randomized trials for unresectable hepatocellular carcinoma: Chemoembolization improves survival.肝脏病学37岁,429 - 442。doi: 10.1053 / jhep.2003.50047
Llovet, j . M。,Kelley, R. K., Villanueva, A., Singal, A. G., Pikarsky, E., Roayaie, S., et al. (2021). Hepatocellular carcinoma.Nat。启说,拘谨的。7、6。doi: 10.1038 / s41572 - 020 - 00240 - 3
洛瑞,o . H。,Rosebrough, N. J., Farr, A. L., and Randall, R. J. (1951). Protein measurement with the Folin phenol reagent.生物。化学。193年,265 - 275。doi: 10.1016 / s0021 - 9258 (19) 52451 - 6
谅解备忘录,Y。,Wang, J., Wu, J., Hu, D., Zhang, C., Duan, C., et al. (2019). Ferroptosis, a new form of cell death:opportunities and challenges in cancer.j .内科杂志。肿瘤防治杂志。12,34-50。doi: 10.1186 / s13045 - 019 - 0720 - y
Ng, i O。,Liu, C. L., Fan, S. T., and Ng, M. (2000). Expression of P-glycoprotein in hepatocellular carcinoma: A determinant of chemotherapy response.点。j .中国。病理学研究。113年,355 - 363。doi: 10.1309 / AC1M-4TY4-U0TN-EN7T
Ogunwobi, O . O。,Harricharran, T., Huaman, J., Galuza, A., Odumuwagun, O., Tan, Y., et al. (2019). Mechanisms of hepatocellular carcinoma progression.世界j .杂志。25日,2279 - 2293。doi: 10.3748 / wjg.v25.i19.2279
公园,W。,Chung, Y. H., Kim, J. A., Jin, Y. J., Lee, D., Shim, J. H., et al. (2013). Recurrences of hepatocellular carcinoma following complete remission by transarterial chemoembolization or radiofrequency therapy: Focused on the recurrence patterns.乙醇。Res。43岁,1304 - 1312。doi: 10.1111 / hepr.12083
Pozarowski, P。,Halicka, D. H., and Darzynkiewicz, Z. (2003). NF-kappaB inhibitor sesquiterpene parthenolide induces concurrently atypical apoptosis and cell necrosis: Difficulties in identification of dead cells in such cultures.Cytom。一个54岁,118 - 124。doi: 10.1002 / cyto.a.10057
Ralstin, m . C。计,大肠。,Yip-Schneider, M. T., Klein, P. J., Wiebke, E. A., and Schmidt, C. M. (2006). Parthenolide cooperates with NS398 to inhibit growth of human hepatocellular carcinoma cells through effects on apoptosis and G0-G1 cell cycle arrest.摩尔。癌症Res。4,387 - 399。doi: 10.1158 / 1541 - 7786. - mcr - 05 - 0157
西格尔,r . L。,Miller, K. D., and Jemal, A. (2019). Cancer statistics, 2019.Ca。j .中国癌症。69年,7-34。doi: 10.3322 / caac.21551
Sobota, R。,Szwed, M., Kasza, A., Bugno, M., and Kordula, T. (2000). Parthenolide inhibits activation of signal transducers and activators of transcription (STATs) induced by cytokines of the IL-6 family.物化学。Biophys。Commun >,267年,329 - 333。doi: 10.1006 / bbrc.1999.1948
斯塔克。,Wang, C., Savic, L. J., Letzen, B., Schobert, I., Miszczuk, M., et al. (2020). Automated feature quantification of Lipiodol as imaging biomarker to predict therapeutic efficacy of conventional transarterial chemoembolization of liver cancer.科学。代表。10日,18026年。doi: 10.1038 / s41598 - 020 - 75120 - 7
太阳,J。,Zhang, C., Bao, Y.-L., Wu, Y., Chen, Z.-L., Yu, C.-L., et al. (2014). Parthenolide-induced apoptosis, autophagy and suppression of proliferation in HepG2 cells.亚洲pac。j .癌症上一页。15日,4897 - 4902。doi: 10.7314 / apjcp.2014.15.12.4897
太阳,Y。,Clair, D. K. S., Xu, Y., Crooks, P. A., and Clair, W. H. S. (2010). A NADPH oxidase–dependent redox signaling pathway mediates the selective radiosensitization effect of parthenolide in prostate cancer cells.癌症Res。70年,2880 - 2890。doi: 10.1158 / 0008 - 5472. -能- 09 - 4572
Ursini F。,Maiorino, M., Hochstein, P., and Ernster, L. (1989). Microsomal lipid peroxidation: Mechanisms of initiation: The role of iron and iron chelators.自由·拉迪奇。医学杂志。地中海。6,31-36。0891 - 5849 . doi: 10.1016 / (89) 90156 - 1
温,J。,You, K.-R., Lee, S.-Y., Song, C.-H., and Kim, D.-G. (2002). Oxidative stress-mediated apoptosis: The anticancer effect of the sesquiterpene lactone parthenolide.生物。化学。277年,38954 - 38964。doi: 10.1074 / jbc.M203842200
徐,Y。,Fang, F., Miriyala, S., Crooks, P. A., Oberley, T. D., Chaiswing, L., et al. (2013). KEAP1 is a redox sensitive target that arbitrates the opposing radiosensitive effects of parthenolide in normal and cancer cells.癌症Res。73年,4406 - 4417。doi: 10.1158 / 0008 - 5472. - 12 - 4297
Yu H。,Guo, P., Xie, X., Wang, Y., and Chen, G. (2016). Ferroptosis, a new form of cell death, and its relationships with tumourous diseases.j .细胞。摩尔。地中海。21 (4),648 - 657。doi: 10.1111 / jcmm.13008
张,S。,Ong, C.-N., and Shen, H.-M. (2004). Critical roles of intracellular thiols and calcium in parthenolide-induced apoptosis in human colorectal cancer cells.癌症。208年,143 - 153。doi: 10.1016 / j.canlet.2003.11.028
张,Y。,Zhang, M., Chen, M., Mei, J., Xu, L., Guo, R., et al. (2018). Association of sustained response duration with survival after conventional transarterial chemoembolization in patients with hepatocellular carcinoma.JAMA Netw。开放1,e183213。doi: 10.1001 / jamanetworkopen.2018.3213
关键词:肝细胞癌(HCC), parthenolide (PTL),硫醇氧化、脂质过氧化反应,ferroptosis,谷胱甘肽
引用:LoBianco阵线,Krager KJ,约翰逊E,古德温有限公司艾伦AR,骗子PA,密友厘米,保华乔丹和Aykin-Burns N (2022) Parthenolide诱发快速硫醇氧化导致ferroptosis肝癌细胞。前面。Toxicol。4:936149。doi: 10.3389 / ftox.2022.936149
收到:2022年5月04;接受:2022年11月07;
发表:2022年12月14日。
编辑:
Stina Oredsson瑞典隆德大学审核:
Pasupathi Sundaramoorthy美国卡罗莱纳州BioOncology研究所(CBI)智史暨南大学,中国
Ranjana Kumari Kanchan美国内布拉斯加州大学医学中心
版权©2022 LoBianco Krager,约翰逊,古德温,艾伦,骗子,密友,保华和Aykin-Burns。这是一个开放分布式根据文章知识共享归属许可(CC)。使用、分发或复制在其他论坛是允许的,提供了原始作者(年代)和著作权人(s)认为,最初发表在这个期刊引用,按照公认的学术实践。没有使用、分发或复制是不符合这些条件的允许。
*通信:Nukhet Aykin-Burns,NAykinBurns@uams.edu